Using this protocol all live secondary stem cell and tissue types have been shown to be susceptible to A. tumefaciens transformations and have been defined into sector types based on the cell type initially transformed and it subsequent developmental growth pattern. Sector types include periderm, phloem, cambial, wound parenchyma and tylose (Figure 1b, 1c, 1d) and can be found in consistent locations describe in the remainder of this paragraph. A periderm sector comprises of a group of transformed cells found exclusively in the periderm and never extending into the phloem (Figure 1e). A phloem sector can be found at various locations between the initiating layer and the periderm, however, never extending into these surrounding tissues (Figure 1f). A cambial sector is comprised of transformed xylem/cambium/phloem tissues derived from transformation of cambial initial and can occur in three distinct patterns: i) 'full' cambial sector, transformed xylem/cambial/phloem cells extending from the boundary of the wound parenchyma tissue through the xylem and cambial zone to the phloem tissue containing both ray and fusiform cells or ray elements only (Figure 1g), ii) 'lost initial' cambial sector, a variant of the full cambial sector where an initial has been lost from the initiating layer leaving mirrored xylem and phloem sectors and an untransformed initiating layer (Figure 1h), and iii) 'xylem only' cambial sector, transformed xylem tissue extending from the boundary of the wound parenchyma tissue to variable lengths into the newly formed xylogenic tissue but never reaching the initiating layer (Figure 1i). A wound parenchyma sector contains transformed wound parenchyma cells found as either an individual cell or groups of cells, often in radial files, located between the pre-existing wood and newly formed xylem tissue (Figure 1j). A tylose Sector comprised of transformed vessel tyloses found within the pre-existing wood (Figure 1k).
Representative transformation efficiency data for both gene and promoter of interest studies as well as their positive controls is presented in Table 1 and Table 2. Data has been drawn from two large trials conducted across the same time period (Dec-Apr) in the same genotypes of eucalypt and poplar across two successive years involving a number of wood formation genes and promoters of interest. Focusing on positive controls from the promoter of interest studies (Table 2), the most susceptible tissue type to A. tumefaciens T-DNA transfer is cambial tissues with approximately 60% and 40% of the sectors identified in eucalypts and poplar respectively being cambial sectors. In eucalypts, the next most abundant sector type is wound parenchyma at 35% while the other sector types all had ATScm-2 values below 5%. In poplars, the next most abundant sector type is wound parenchyma at 20% followed by phloem at 15% while the remainder of the sector types were found at a frequency below 5%. A higher likelihood of finding a sector in a cambial window as well as greater numbers of cambial sectors identified are typical where the phloem peel protocol is used (Table 1, step 5.2.1) due to the ability for staining and the possibility to visualize the entire area of the cambium.
During the development of these protocols a number of variables were explored as part of protocol optimization trials. These included bacterial concentration in Inoculation Media, media type used in Inoculation Media, addition of Acetosyringone to Inoculation Media, tissue age and A. tumefaciens strain as well as time of inoculation and genotype. Cambial sector ATScm-2 data from these trials is presented in Table 3 and Table 4. The majority of variables investigated, when altered, lead to changes in cambial ATScm-2 in both species and included in particular increasing the concentration of bacteria in the Inoculation Media, use of MS in the Inoculation Media and choice of A. tumefaciens strain. In addition, time of inoculation and tree genotype showed significant differences in eucalypts with inoculations in early summer showing higher ATScm-2 while eucalypt clones SG21 and SG44 showed higher cambial ATScm-2, 41.6 and 40.4, respectively compared to others across all months combined.
Average cambial sector size varies and is dependent on the amount of its tangential and radial growth in a cambial window. In one subsample of 53 sectors in poplar, grown for approximately 4 months, the tangential width of sectors at the cambium varied between 0.09 and 0.58 mm with an average of 0.27 mm while radial growth across cambial windows ranged from 0.7 to 2.2 mm with an average of 1.35 mm. When combined, these provided between 0.063 mm2 and 1.276 mm2 of transgenic tissue for analysis. In another subsample of 188 sectors in the same species, where between 1.5-4 mm of radial growth across the cambial window was observed over approximately 5 months, the average sector mass was 72 µg (Table 5).
The amount of transgenic tissue created has been shown to provide sufficient cells and/or tissue to undertake morphological measurements as well as for the study of promoter activity. For example, the influence of three Eucalyptus grandis FLAs on a number of xylem fiber morphological traits9 was explored in eucalypt clones and revealed possible roles for EgrFLA2 in MFA determination and EgrFLA1 in cell size determination (Table 6). In addition, patterns of expression of Eucalyptus grandis CESA gene promoter in the developing xylem (X1), developed xylem (X2) and phloem (P) tissues10 identified significantly different expression patterns between EgrCESA1, 2, 3, and EgrCESA4, 5, 7 in eucalypt stems. In this analysis, EgrCESA1, 2, 3 were shown to be primarily expressed in developing xylem tissue while EgrCESA4, 5, 7 were shown to be expressed in both developing xylem and phloem tissue (Figure 3b, Table 7). All EgrCESA promoters showed significantly different expression patterns to the positive control (CAMV35S promoter) (Table 7).
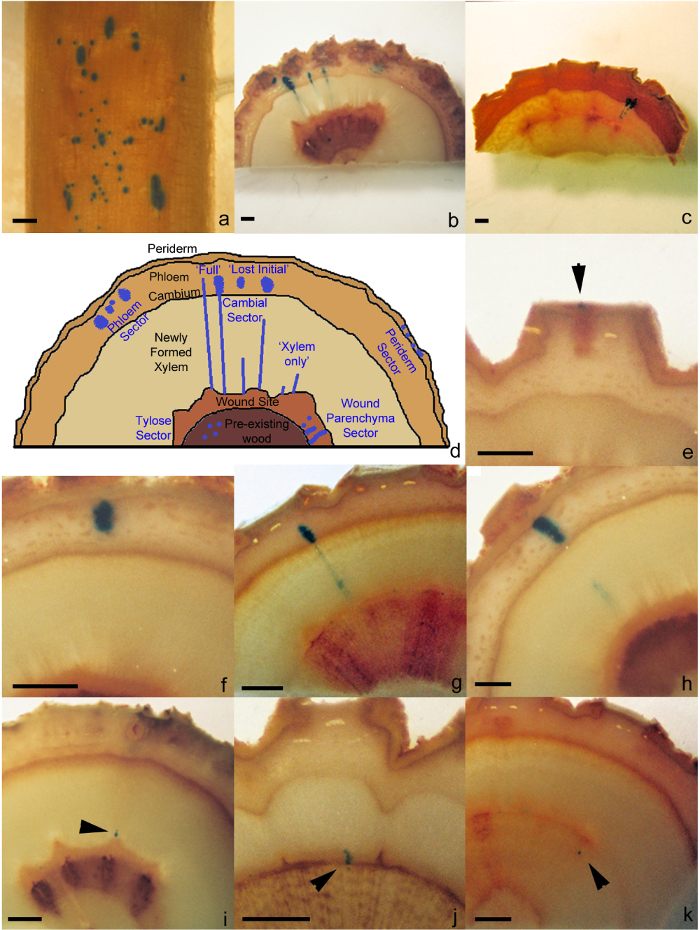
Figure 1: Transgenic sectors types. (a) Cambial sectors as viewed on the surface of the exposed xylem after processing using the phloem peel protocol (step 5.2.1). Discs showing a range of sector types on transverse surface of in poplar (b) and eucalypt (c) stems after processing using the transverse cut protocol (step 5.2.2). (d) Schematic diagram of the range of sector types found in plant stems. Examples of periderm sector (e), phloem sector (f), 'full' cambial sector (g), 'lost initial' cambial sector (h), 'xylem only' cambial sector (i), wound parenchyma sector (j) and tylose sector (k) in poplar. Black arrows indicate where smaller sized sector are located. All scale bars = 1 mm. Please click here to view a larger version of this figure.

Figure 2: Preparation of cambial sectors for morphological analysis using SEM and light microscopy. (a) Excision of disc containing sector. (b) Disc trimmed to remove excess tissue. Cutting through transverse surface of the transgenic sector (c). (d) Introduction of two radial cuts to assist in locating the transgenic tissue under SEM. (e) Tissue mounted on an SEM pin using conductive tape. (f) Depiction of the region of transgenic and non-transgenic tissues that should be imaged for analysis. (g) Typical image of xylem fiber and ray cells captured at 2,000X magnification. (h) Area where transgenic and non-transgenic fibers should be excised from for MFA analysis. (i) Macerated fiber viewed under light microscopy depicting the angle of the pits and/or cell wall striations and to the long axis of the cell. Black arrow indicates pit aperture. Scale bars = a-f, h = 1 mm, g, i = 20 µm. Please click here to view a larger version of this figure.
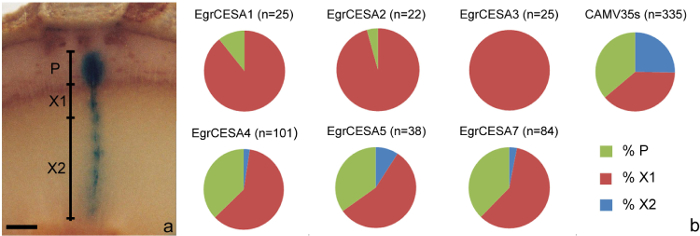
Figure 3: Analysis and results of gene promoter expression patterns of cambial derivatives. (a) Depiction of the P (phloem), X1 (developing xylem) and X2 (developed xylem) regions. (b) Results indicating proportion of P, X1 and X2 staining in cambial sectors from a trial involving a range of CESA promoters from Eucalyptus grandis transformed into stems of eucalypt hybrids. Data sourced from Creux et al.10. n = number of sectors assessed. Scale bar = 0.5 mm. Please click here to view a larger version of this figure.
| Tree species | Vector type | Number of genes of interest used | Total number of windows created | Total number of windows where a sector was identified (%) | Cambial ATScm-2 cambial (in windows where a sector was identified) |
| Eucalyptus globulus x camaldulensis hybrids (total of three clones) | Positive Control (Genes of interest) | 1 (GUS only) | 96 | 97% | 45.1 (1.5) |
| Genes of Interest | 20 | 630 | 92% | 46.1 (2.2) | |
| Populus alba 'pyramidalis' | Positive Control (Genes of interest) | 1 (GUS only) | 110 | 74% | 10.8 (1.5) |
| Genes of Interest | 20 | 700 | 81% | 14.6 (0.8) |
Table 1: Typical cambial ATScm-2 for the positive control and genes of interest. Figures have been sourced from studies undertaken over two consecutive years in the same eucalypt and poplar clones using a range of genes and the phloem peel harvesting protocol (step 5.2.1). Standard error values in brackets.
| Tree species | Vector type | Number of promoter/genes | Total number of windows created | Total number of windows where a sector was identified (%) | ATScm-2 all sectors | ATScm-2 Periderm | ATScm-2 Phloem | ATScm-2 cambial | ATScm-2 Wound Parenchyma | ATScm-2 Tylose |
| Eucalyptus globulus x camal- dulensis hybrids (total of three clones) |
Positive Control | 1 (GUS only) | 118 | 63% | 25.74 (2.35) | 0.24 (0.07) | 0.89 (0.19) | 15.7 (1.71) | 8.84 (1.4) | 0.07 (0.03) |
| Promoters of Interest | 28 | 1050 | 31% | 14.67 (1.77) | 0.02 (0.01) | 0.05 (0.01) | 13.79 (1.75) | 0.78 (0.21) | 0.03 (0.01) | |
| Populus alba 'pyramidalis' | Positive Control | 1 (GUS only) | 110 | 54% | 12.37 (1.77) | 0.22 (0.07) | 1.85 (0.29) | 5.07 (0.6) | 2.78 (0.75) | 0.29 (0.53) |
| Promoters of Interest | 28 | 990 | 26% | 8.28 (1.18) | 0.16 (0.04) | 0.9 (0.09) | 6.67 (1.16) | 0.27 (0.07) | 0.28 (0.11) |
Table 2: Typical sector ATScm-2 for the positive control and promoters of interest. Figures have been sourced from studies undertaken over two consecutive years in the same eucalypt and poplar clones using a range of promoter sequences and the disc harvesting protocol (step 5.2.2). ATScm-2 values were derived from windows where a sector was identified only. Standard error values in brackets. Please click here to view a larger version of this table.
| Species | Trial | Treatment Description | Number of windows in treatment | Cambial ATScm-2 |
| Eucalyptus globulus | Concentration of A. tumefaciens in Inoculation Media | Resuspended in 25 ml MS | 10 | 1.4 (0.62) |
| Resuspended in 5 ml MS | 10 | 1.6 (0.72) | ||
| Resuspended in 1 ml MS (control) | 10 | 2.4 (1.16) | ||
| Media type in Inoculation Media | LB | 10 | 0.4 (0.22) | |
| MS (control) | 10 | 2.4 (1.16) | ||
| Addition of Acetosyringone to Inoculation Media | With Acetosyringone | 11 | 0.9 (0.37) | |
| Without Acetosyringone (control) | 11 | 0.6 (0.64) | ||
| Age of stem tissue inoculated | 6 months (control) | 11 | 1.1 (0.66) | |
| 18 months | 11 | 1.8 (0.95) | ||
| A. tumefaciens strain | AGL1 (control) | 18 | 0 (0) | |
| C58 | 18 | 0.1 (0.1) | ||
| LBA4404 | 18 | 0.6 (0.4) | ||
| Populus alba 'pyramidalis' | Concentration of A. tumefaciens in media | Resuspended in 25 ml MS | 10 | 2.2 (0.55) |
| Resuspended in 5 ml MS | 10 | 2.7 (0.63) | ||
| Resuspended in 1 ml MS (control) | 10 | 5.1 (1.58) | ||
| Media type used for stem inoculation | LB | 10 | 2.8 (0.71) | |
| MS (control) | 10 | 5.1 (1.58) | ||
| Addition of Acetosyringone to Inoculation Media | With Acetosyringone | 11 | 0.3 (0.14) | |
| Without Acetosyringone (control) | 11 | 0.5 (0.25) | ||
| Age of stem tissue inoculated | 6 months (control) | 11 | 1.5 (0.33) | |
| 18 months | 11 | 1.5 (0.63) | ||
| A. tumefaciens strain | AGL1 (control) | 20 | 8.1 (2) | |
| C58 | 20 | 12.5 (2) | ||
| LBA4404 | 20 | 11.1 (2) |
Table 3: Cambial ATScm-2 values for a range of variables investigated as part of protocol optimization trails. Variables were investigated in a poplar clone and a range of Eucalyptus globulus individuals across a number of years with cambial ATScm-2 data presented. Cambial windows were harvested using the disc harvesting protocol (step 5.2.2). Standard error values in brackets.
| Eucalyptus globulus x camaldulensis clone ID | Number of cambial windows for each month | Cambial ATScm-2 Late Spring (November) Inoculation | Cambial ATScm-2 Early Summer (December) Inoculation | Cambial ATScm-2 Mid Summer (January) Inoculation | Cambial ATScm-2 Late Summer (February) Inoculation | Cambial ATScm-2 Early Autumn (March) Inoculation | Cambial ATScm-2 all months combined |
| SG5 | 10 | 15.7 (7.8) | 50.7 (13.7) | 10.5 (5.1) | 30.7 (4.4) | 16.8 (6.3) | 24.9 (4.3) |
| SG6 | 10 | 6.1 (3.1) | 38.5 (15.1) | 4.5 (1.7) | 14.5 (3.3) | 27.6 (8.4) | 18.3 (4) |
| SG13 | 10 | 28.4 (6.7) | 41.8 (9.1) | 16.1 (7) | 29.3 (5.2) | 19.8 (4.3) | 27.1 (3.1) |
| SG18 | 10 | 6.7 (2.9) | 18.3 (6.7) | 17.4 (3.4) | 19.2 (6.1) | 18.1 (6.3) | 15.5 (2.4) |
| SG21 | 10 | 38.7 (14.3) | 77 (17.5) | 23.9 (5.7) | 27.5 (6.9) | 40.7 (13.5) | 41.6 (6) |
| SG35 | 10 | 23.5 (10.2) | 42.8 (12.2) | 7.6 (2.5) | 17.6 (4.3) | 31.3 (4.6) | 24.6 (3.8) |
| SG37 | 10 | 19.7 (8) | 55.2 (14.5) | 7.4 (1.8) | 35 (10.4) | 15.9 (4.5) | 26.6 (3.8) |
| SG39 | 10 | 12.5 (2.4) | 23.6 (6.3) | 6.9 (3) | 26.3 (8.1) | 7.3 (2.3) | 15.5 (2.6) |
| SG40 | 10 | 24.8 (11) | 24.5 (6.8) | 14 (5.6) | 35.6 (6.1) | 19.9 (4.2) | 23.8 (3.2) |
| SG44 | 10 | 22.3 (3.4) | 63 (29.3) | 9.8 (2.6) | 52.5 (10.3) | 54.3 (15) | 40.4 (7.4) |
| SG46 | 10 | 15.3 (5.2) | 63.9 (20.1) | 23.6 (4) | 22.5 (4.8) | 17.4 (4.9) | 28.5 (5.1) |
| Monthly Cambial ATScm-2 | 19.9 (5) | 45.3 (9.1) | 12.6 (2.8) | 27.8 (4.5) | 24 (5.1) |
Table 4: Influence of genotype and time of inoculation on cambial ATScm-2 in eucalypt clones. Ten windows were introduced each month over the growing season to 10 different Eucalyptus globulus x camaldulensis clones which were all harvested in May. Monthly data is presented for each genotype with an overall average monthly and genotypic ATScm-2 presented also. Cambial windows were harvested using the phloem peel harvesting protocol (step 5.2.1). Standard error values in brackets. Please click here to view a larger version of this table.
| Plant Number | Number of cambial sectors excised | Total mass of all sectors (µg) | Average mass of sectors (µg) |
| 1 | 11 | 750 | 68 |
| 2 | 19 | 1,460 | 77 |
| 3 | 21 | 160 | 8 |
| 4 | 26 | 4,460 | 172 |
| 5 | 15 | 1,320 | 88 |
| 6 | 22 | 2,490 | 113 |
| 7 | 19 | 1,720 | 91 |
| 8 | 30 | 830 | 28 |
| 9 | 25 | 360 | 14 |
| Total | 188 | 13,550 | 72 |
Table 5: Typical mass of a cambial sector. 45 windows across nine plants were investigated in poplar to determine the average mass of a sector and harvested using the disc harvesting protocol (step 5.2.2). Plants were grown for 5 months and the radial growth across a cambial window was between 1-4 mm.
| Morph- ological trait |
GUS only (positive control) | Non-transgenic tissue (GUS only) | P-value | Gene of Interest 1 (FLA1) | Non-transgenic tissue (FLA1) | P-value | Gene of Interest 2 (FLA2) | Non-transgenic tissue (FLA2) | P-value | Gene of Interest 3 (FLA3) | Non-transgenic tissue (FLA3) | P-value |
| Average Cell Wall Thickness (µm)* | 1.81 (0.1) | 1.65 (0.1) | 0.2692 | 1.47 (0.14) | 1.55 (0.12) | 0.6403 | 1.65 (0.13) | 1.78 (0.13) | 0.4839 | 1.59 (0.12) | 1.63 (0.11) | 0.8254 |
| Average Cell Wall Area (µm2)* | 69.1 (4.74) | 60.7 (2.03) | 0.1229 | 64.1 (15.68) | 59.9 (9.79) | 0.5004 | 61.6 (3.4) | 65 (3.96) | 0.5182 | 61.9 (3.56) | 61.5 (3.16) | 0.9438 |
| Average Cell Area (µm2)* | 115.9 (11.01) | 99.9 (5.1) | 0.2041 | 124.4 (6.1) | 108 (4.36) | 0.0445 | 110.8 (8.45) | 111.3 (9.06) | 0.9671 | 108.5 (4.43) | 103 (3.07) | 0.3204 |
| Average Lumen Area (µm2)* | 46.9 (7.55) | 39.2 (4.89) | 0.407 | 60.2 (5.28) | 48.1 (4.46) | 0.0991 | 49.2 (7.56) | 46.3 (7.5) | 0.7882 | 46.6 (3.31) | 41.5 (3.9) | 0.3264 |
| Average Microfibril Angle (O)# | 24.3 (0.75) | 24.2 (0.45) | 0.8648 | 22.3 (0.82) | 22.7 (0.7) | 0.5664 | 23.7 (0.55) | 26.6 (0.5) | 0.0001 | 26 (0.88) | 27.2 (0.72) | 0.0903 |
Table 6: ISSA Fiber morphology measurements derived from gene of interest study. Comparison of fiber cell wall, cell size and MFA measurements taken from transgenic fibers sourced from Eucalyptus globulus x camaldulensis clones stems transformed with Eucalyptus grandis FLAs (EgrFLA1, 2, 3) and positive control (GUS only) with the adjacent non transgenic control fibers. Data sourced from MacMillan et al.9. * indicates trial involved the measurement of 10 fibers in 10 sector (total of 100 fibers). # indicates trial involved the measurement of 5 fibers in 20 sectors (total of 100 fibers). α <0.05 shown in bold. Please click here to view a larger version of this table.
| EgCESA1 | EgCESA2 | EgCESA3 | EgCESA4 | EgCESA5 | EgCESA7 | GUS only (positive control) | |
| EgrCESA1 | 0.708 | 2.839 | 8.856 | 9.976 | 9.18 | 29.165 | |
| EgrCESA2 | 1.11 | 11.05 | 12.008 | 11.363 | 30.234 | ||
| EgrCESA3 | 15.151 | 16.123 | 15.488 | 37.791 | |||
| EgrCESA4 | 4.852 | 0.108 | 46.858 | ||||
| EgrCESA5 | 3.275 | 11.278 | |||||
| EgrCESA7 | 36.082 | ||||||
| GUS only (positive control) |
Table 7: Chi2 values derived from comparison of promoter expression patterns in cambial derivatives. Chi2 analysis comparing the presence/absence of GUS staining in P, X1 and X2 region of cambial derivatives between six Eucalyptus grandis CESA gene promoters sequences (EgrCESA 1, 2, 3, 4, 5, 7) and positive control (GUS only). Data sourced from Creux et al.10. α <0.05 shown in underline.
