1. Chemicals
- Prepare a solution of 500 mM CEPA (144.49 g/mol), and a solution consisting of both 500 mM NaCl (58.44 g/mol) and 500 mM NaH2PO4·H2O (137.99 g/mol) in acidified (pH 2.5) ultra-pure water or 0.1 N HCl. Mix using a vortex until the solutions are clear.
- Serially dilute (10x) the 500 mM solutions in the same solvent to obtain 5 mM and 50 mM stocks.
- Prepare a 10 mM solution of 1-aminocyclopropane carboxylic acid (ACC; 101.1 g/mol) in ultra-pure water.
- Filter-sterilize stock solutions and store aliquots at -20 °C.
- Filter-sterilize cellulase. Store aliquots at 4 °C.
2. Verifying Ethylene Production from 2-Chloroethylphosphonic Acid Decomposition: Triple Response Assay
- Surface-sterilize Arabidopsis thaliana Ecotype Columbia Seeds Using the Vapor-phase Method:
CAUTION: The following step produces toxic chlorine gas. Conduct seed sterilization in a fume hood.- Obtain a sealable container deep enough to accommodate a 250 ml glass beaker for seed sterilization and place it in a fume hood.
- Add A. thaliana seeds to a microcentrifuge tube and place the tube in a rack. Place the rack containing the open tube into the sealable container.
Note: Do not fill individual tubes over half full to allow chlorine gas to penetrate the lower seeds. - Place a 250 ml beaker containing 100 ml of commercial bleach in the sealable container. Carefully add 3 ml of concentrated HCl to the bleach and immediately seal the container with its lid.
CAUTION: Bleach and HCl react to produce toxic chlorine gas which acts to surface-sterilize the seeds. - Incubate the seeds in the presence of chlorine gas for 4 hr in the fumehood.
Note: Longer sterilization times will reduce germination efficiency. - After sterilization, allow the chlorine gas to vent in the fume hood for at least 1 hr then seal the microcentrifuge tube. Leave the bleach and HCl mixture in the fumehood for at least 24 hr before discarding.
Note: Sterilized seeds may be stored at room temperature for immediate use. - Keep seeds at 4 °C for long-term storage. Bring seeds to room temperature before opening the microcentrifuge tube to prevent condensation. Store seeds in the dark.
- Prepare Agar Plates in 4-sector Petri Dishes (90 x 15 mm):
- Prepare 110 ml of growth medium for A. thaliana seedlings: 1x Murashige and Skoog (MS) basal medium15 (4.33 g/L) containing 1% (w/v) sucrose and 0.8% (w/v) agar. Adjust the MS medium to pH 6 with NaOH.
- Prepare 100 ml of bacterial growth medium for CEPA decomposition: Schramm and Hestrin (SH) medium16 containing 1.5% (w/v) agar. Adjust the SH medium to pH 7 with NaOH.
- Sterilize media by autoclaving and temper in a 55 °C water bath.
- Once the MS agar has tempered, add 40 ml to a sterile flask. To prepare the positive control growth medium for A. thaliana seedlings, supplement the 40 ml aliquot of MS agar with 40 µl of 10 mM ACC to obtain a final concentration of 10 µM ACC.
- Add 5 ml of medium to the appropriate quadrants of the sectored Petri dishes (Figure 1) and allow the agar to solidify. Prepare all plates in triplicate.
Note: CEPA is not added to the growth medium until later in the protocol.

Figure 1: Setup of agar plates used for the triple response assay with CEPA. A schematic illustrates the quadrants specific for the negative control (A), positive control (B), and experimental plates (C). This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.
- Stratify A. thaliana Seeds to Ensure Synchronous Germination:
- Add approximately fifty A. thaliana seeds onto each quadrant containing MS or MS+ACC agar. Ensure seeds are evenly distributed to facilitate seedling removal and analysis.
- Incubate plates containing seeds in the dark at 4 °C for 3-4 days.
- Decompose CEPA on Bacterial Growth Medium (SH) and Perform Triple Response Assay:
- After stratification, expose the seeds to fluorescent light for 2 hr.
- Spread 10 µl of the 500 mM CEPA stock solution onto the quadrants of the experimental plates containing SH agar (pH 7; Figure 1) to obtain a final CEPA concentration of 1 mM.
- Seal the plates with laboratory film and cover with foil to create a dark environment for the seeds.
- Germinate seeds by incubating plates in the dark at 23 °C for 3 days with the agar side down.
Note: Plates can be stacked, but negative controls should be placed on the bottom since ethylene is lighter than air.
- Analyze Triple Response Assay Data:
- With flame-sterilized forceps, remove single seedlings from a plate corresponding to each treatment and view under a dissecting or digital USB microscope. Ensure the bottom of the seedlings are aligned and photograph.
- Remove 30 seedlings from each quadrant (60 seedlings per biological replicate and 180 seedlings per treatment). Align on a surface with a black background and photograph with a ruler.
- Measure the hypocotyl length (mm) of replicate seedlings using ImageJ software17. Set the scale by clicking and dragging the *Straight* tool to select a length of 10 mm. Select "Set Scale" under the "Analyze" tab and set the known distance to 10. Using the *Segmented* tool, click and drag to select the length of the hypocotyl and press M to measure distance. Compare the means of biological replicates using a one-way ANOVA with a Tukey's multiple comparison test. Differences are considered significant if p < 0.05.
3. Analysis of pH throughout Bacterial Growth
- Grow and Quantify Komagataeibacter xylinus Starter Cultures in Triplicate:
- Inoculate a single colony of K. xylinus into 5 ml of SH medium (pH 5) supplemented with 0.2% (v/v) filter-sterilized cellulase. Incubate cultures at 30 °C with agitation at 150 rpm until an OD600 of 0.3 to 0.4 is reached (about 72 hr).
- Harvest starter cultures by centrifugation (2,000 x g; 4 °C; 10 min). With 5 ml sterile 0.85% (w/v) NaCl solution, wash the cells twice and resuspend the cell pellet. Keep cells on ice.
- Quantify cells using a Petroff-Hausser counting chamber.
- Inoculate Cultures for Analysis of pH:
- Add 150 ml of SH broth (pH 7) supplemented with 0.2% (v/v) filter-sterilized cellulase to twelve 500 ml flasks with foil lids. Inoculate flasks with K. xylinus starter cultures at a concentration of 105 cells/ml. Using the three starter cultures, prepare three biological replicates per treatment.
- Supplement cultures with 300 µl of the 5, 50, or 500 mM CEPA stock solutions to obtain final CEPA concentrations of 0.01, 0.1, and 1.0 mM, respectively. Supplement the untreated control cultures with 300 µl of the solvent used to dissolve CEPA.
- Seal the flasks by tightly taping the foil lids to the flasks and incubate cultures for 14 days at 30 °C with agitation at 150 rpm.
- Each day, aseptically remove 5 ml samples from each flask and pellet cells by centrifugation (2,000 x g; 4 °C; 10 min). Transfer supernatants to a clean tube and measure the pH of each biological replicate using a pH meter.
- Analyze the time-course data by graphing the mean pH of the biological replicates.
Note: It is important that the culture pH does not drop below 3.5; a culture pH of 5 significantly reduces ethylene release from CEPA, and a pH below 3.5 completely inhibits ethylene release. - To control for chlorine and phosphate levels, perform an identical experiment using 0.01, 0.1, and 1.0 mM of the NaCl-NaH2PO4 solution using the 5, 50, and 500 mM stocks, respectively.
4. Colony Morphology
- Grow K. xylinus Starter Cultures in Triplicate:
- Inoculate a single colony of K. xylinus into 5 ml of SH medium (pH 5) supplemented with 0.2% (v/v) filter-sterilized cellulase. Incubate cultures at 30 °C with agitation at 150 rpm until an OD600 of 0.3 to 0.4 is reached (about 72 hr).
- Harvest starter cultures by centrifugation (2,000 x g; 4 °C; 10 min). With 5 ml of sterile saline, wash the cells and then resuspend the cell pellet. Keep cells on ice.
- Prepare 24 agar plates containing 25 ml of SH (pH 7) medium with 1.5% (w/v) agar.
- Once the agar has solidified, spread 50 µl of the 5, 50, and 500 mM CEPA stock solutions on the agar to obtain final CEPA concentrations of 0.01, 0.1, and 1.0 mM, respectively. Set up untreated and solvent control plates, which consist of no amendment and spreading of 50 µl of the solvent used to dissolve the test compounds.
- Streak plates for isolated colonies with a loop-full (~5 μl) of K. xylinus starter culture and seal them with paraffin film. Inoculate all plates in triplicate. Incubate plates for five days at 30 °C.
- Photograph colonies at 20X magnification using a digital USB microscope and qualitatively assess colonial morphology and cellulose production on solid medium.
Note: Cellulose appears as a hazy substance along the margin of the colony. - To control for chlorine and phosphate levels, perform an identical experiment using 0.01, 0.1, and 1.0 mM of the NaCl-NaH2PO4 solution using the 5, 50, and 500 mM stocks, respectively.
5. Pellicle Assays
- Grow and Quantify K. xylinus Starter Cultures in Triplicate:
- In triplicate, inoculate a single colony of K. xylinus into 5 ml of SH medium (pH 5) supplemented with 0.2% (v/v) filter-sterilized cellulase. Incubate cultures at 30 °C with agitation at 150 rpm until an OD600 of 0.3 to 0.4 is reached (about 72 hr).
- Harvest starter cultures by centrifugation (2,000 x g; 4 °C; 10 min). With 5 ml of sterile saline, wash the cells twice and resuspend the cell pellet. Keep cells on ice.
- Quantify cells using a Petroff-Hausser counting chamber.
- Prepare Master Mixes to Inoculate 24-well Plates:
- Supplement 60 ml of SH medium (pH 7) with 120 µl of the 5, 50, and 500 mM CEPA stocks to obtain final CEPA concentrations of 0.01, 0.1, and 1.0 mM, respectively. Supplement another 60 ml of SH medium (pH 7) with 120 µl of the solvent used to dissolve CEPA. Vortex to mix.
- Divide each 60 ml of CEPA-containing medium into four 14 ml aliquots. Inoculate three of the 14 ml aliquots with biological replicates of starter culture at a concentration of 105 cells/ml. Keep tubes with cells on ice to prevent cellulose production.
Note: The remaining 14 ml aliquot will be used for sterile control wells.
- Inoculate 24-well Plates:
- Test each treatment in its own plate with three rows of biological replicates and a row of sterile controls (Figure 2).
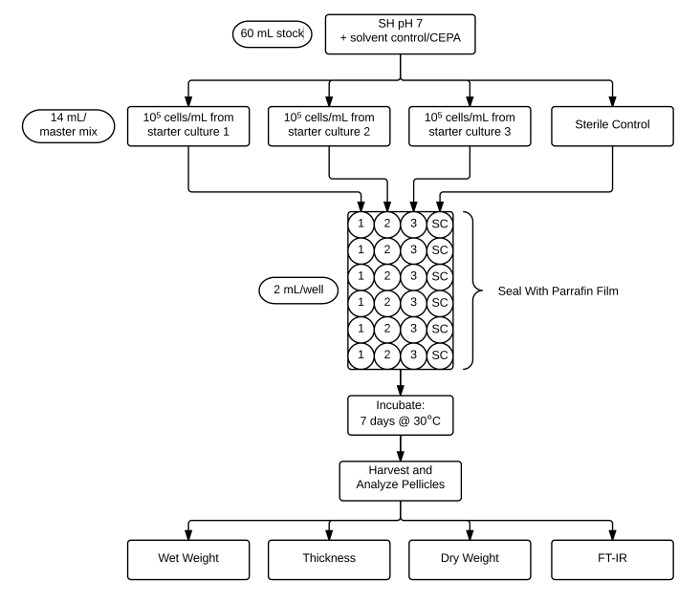
Figure 2: Flow-chart illustrating the protocol used for pellicle assay and analysis. Stock CEPA-supplemented pH 7 SH medium (60 ml) is aliquoted for three separate biological replicate inoculations and a sterile control (14 ml each). These cultures are then aliquoted into six technical replicates (2 ml) into a 24 well plate and then sealed with paraffin film. After incubation for 7 days at 30 °C, pellicles are harvested and characterized by determining wet weight, thickness, dry weight, and crystallinity by FT-IR. Please click here to view a larger version of this figure
- Using the 14 ml master mix, add 2 ml into each of six wells of a sterile 24-well plate. Complete for the three biological replicates and sterile control (Figure 2). Repeat for each treatment.
- Seal plates with paraffin film and incubate statically for 7 days at 30 °C.
- Harvest and Measure Pellicle Wet Weight, Thickness, dry Weight (Cellulose Yield) and Crystallinity (Figure 2):
- Depress one side of the pellicle to elevate the opposing pellicle edge and remove individual pellicles with forceps. While retaining grip, place them on fresh paper towel for 3 sec to remove excess medium before weighing to determine their wet weights.
- Align pellicles adjacent to a ruler and photograph from the side using a high resolution digital camera.
- Using ImageJ software17, measure pellicle thickness on the left shoulder, right shoulder and the center of each pellicle. Average all technical replicates for each biological replicate.
- Individually transfer the pellicles into the wells of a 6-well plate. Treat pellicles with 12 ml of 0.1 N NaOH at 80 °C for 20 min to lyse cells.
- Remove the NaOH and neutralize pellicles by washing with ultra-pure water for 24 hr with agitation. Change water every 6 hr.
Note: Pellicles should be white upon completion of the washing step. - Place pellicles on silicon mats and dry at 50 °C for 48 hr to constant weight. Once dry, remove from mats and measure pellicle weights on an analytical scale to determine bacterial cellulose yield.
- Analyze pellicle crystallinity using Fourier-transform infrared spectroscopy (FT-IR) using 32 scans and a resolution of 4 cm-1 in the range of 4,000 to 650 cm-1. Calculate the crystallinity index, CI(IR), using A1437/A895; the absorbance ratio of the "crystalline band" and "amorphous band" as previously described18.
- To control for chlorine and phosphate levels, perform an identical experiment using 0.01, 0.1, and 1.0 mM of the NaCl-NaH2PO4 solution using the 5, 50, and 500 mM stocks, respectively.
- Analyze Pellicle Data:
- Calculate pellicle hydration by determining the difference between pellicle wet weight and dry weight.
- Average the values of all technical replicates to obtain a single value for each biological replicate for statistical analysis. Compare treatments using a one-way ANOVA with Tukey's multiple comparison test. Differences are significant if p <0.05.
- Normalize data as the percent of untreated controls and plot the means of biological replicates.
A schematic plate setup for verification of ethylene liberation from CEPA in SH medium (pH 7) by the triple response assay is shown in Figure 1A–C. A flow-chart illustrating the pellicle protocol is shown in Figure 2. Dark-grown A. thaliana seedlings exhibit the triple response phenotype (shorter and thicker hypocotyl with an exaggerated apical hook) in the presence of ACC and in the presence of ethylene produced through the decomposition of CEPA on SH medium (pH 7), but not under untreated conditions (Figure 3A)9. The hypocotyl length of ACC- and CEPA-derived ethylene treated A. thaliana seeds were significantly (p < 0.0001) shorter than untreated controls (Figure 3B)9, confirming that ethylene was released from CEPA on SH medium (pH 7) at a physiologically relevant concentration. The pH of untreated and CEPA-treated K. xylinus cultures remained above 5 (Figure 4)9; therefore CEPA decomposition into ethylene was not impaired due to bacterial organic acid production. CEPA-derived ethylene increased bacterial cellulose production when K. xylinus was grown on solid SH medium (pH 7) (Figure 5)9. All concentrations of CEPA-derived ethylene significantly decreased pellicle wet weight (Figure 6A), did not affect pellicle thickness (Figure 6B) and significantly increased pellicle dry weight (cellulose yield; Figure 6C)9. A representative photo taken for pellicle thickness measurement is shown in Figure 7. Pellicle hydration was reduced by all concentrations of CEPA-derived ethylene (Figure 8A)9, while all concentrations of CEPA-derived ethylene9 increased pellicle crystallinity (Figure 8B). The effects of NaCl and NaH2PO4 were insignificant in all cases (data not shown) confirming that observed phenotypes were caused by CEPA-derived ethylene.
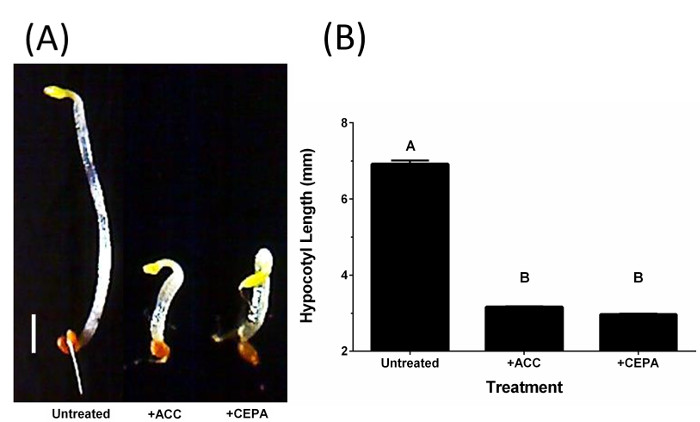
Figure 3: CEPA decomposes on SH medium (pH 7) to produce ethylene. Digital USB microscope photographs show that dark-grown A. thaliana seedlings display the triple response phenotype (shorter hypocotyl, thicker hypocotyl and exaggerated apical hook) when grown in the presence of ACC and CEPA-derived ethylene compared to the untreated control (A). The hypocotyl length of seedlings grown in the presence of ACC and CEPA-derived ethylene were significantly shorter than the untreated control (B). Scale bar = 1 mm. Error bars show SD (n = 3). Bars with different letters are significantly different (p < 0.0001). This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.
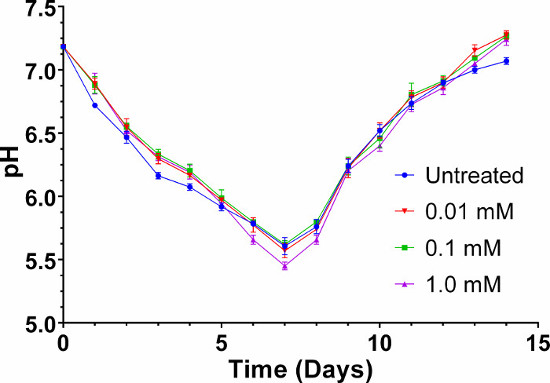
Figure 4: The pH of K. xylinus cultures remains high enough for CEPA decomposition in SH medium (pH 7). The culture pH stays above 5, allowing for efficient decomposition of CEPA into ethylene throughout the bacterial growth cycle. The pH of K. xylinus cultures must be monitored as they secrete and resorb organic acids. The legend shows the CEPA concentrations tested. Note that the y-axis begins at pH 5. Error bars show SD (n = 3). This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.

Figure 5: CEPA-derived ethylene increases cellulose production by K. xylinus on a solid medium. K. xylinus was grown on SH agar plates (pH 7) that were untreated (A), or pre-treated with acidified (pH 2.5) ultra-pure water (B), phosphate and chloride (C–E) or CEPA (F–H). Representative colonies are shown. The arrow shows cellulose produced by K. xylinus, seen as the hazy substance around the colony. Scale bar = 0.5 mm. This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.
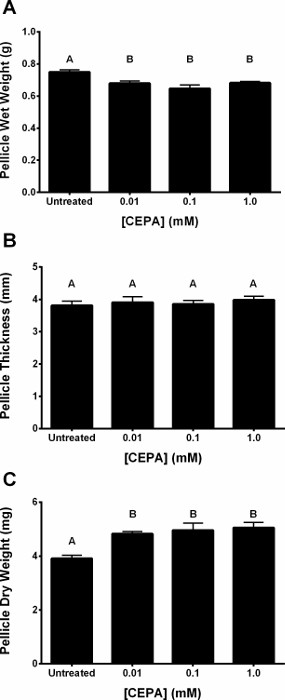
Figure 6: CEPA-derived ethylene decreases the water-holding capacity and increases the yield of K. xylinus cellulose pellicles. Cultures were grown statically in SH broth (pH 7) supplemented with CEPA in 24-well plates, and incubated at 30 °C for 7 days before pellicles were harvested and analyzed. CEPA-derived ethylene decreased pellicle wet weight (A), had no effect on pellicle thickness (B) and increased pellicle dry weight (C). The different CEPA treatments were not significantly different from each other. Error bars show SD (n = 3). Bars with different letters are significantly different (p < 0.05). This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.

Figure 7: Representative photograph of K. xylinus pellicles. Pellicle thickness was measured from photographs using ImageJ software. Scale bar = 10 mm. Please click here to view a larger version of this figure.
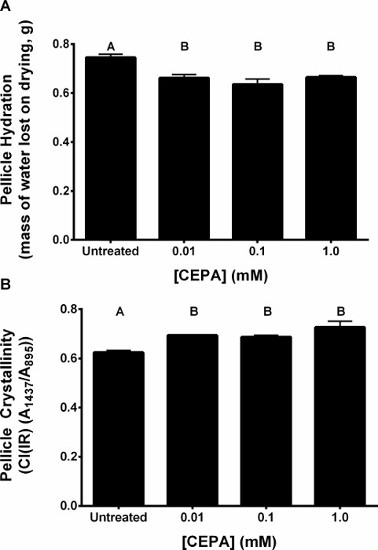
Figure 8: CEPA-derived ethylene reduced the hydration of K. xylinus cellulose pellicles by increasing pellicle crystallinity. Cultures were grown statically in SH broth (pH 7) in 24-well plates, and incubated at 30 °C for 7 days before pellicles were harvested and analyzed. Pellicle hydration was calculated as the difference between pellicle wet and dry weight. CEPA-derived ethylene reduced pellicle hydration (A) by increasing pellicle crystallinity (B). The different CEPA treatments were not significantly different from each other. Error bars show SD (n = 3). Bars with different letters are significantly different (p < 0.05). This figure has been modified from Augimeri and Strap9. Please click here to view a larger version of this figure.
