Studies in this protocol generally follow standardized experimental design and recommended statistical analysis guidelines from the US Environmental Protection Agency (EPA no. 2000.0) for fathead minnows and the Organization for Economic Cooperation and Development (OECD no. 236) for zebrafish. These experimental designs (e.g., increasing replication) can be modified within the current protocol for future studies. Fish culture conditions follow previously published literature17. All experimental procedures and fish culture protocols followed Institutional Animal Care and Use Committee protocols approved at Baylor University.
1. Exposing Fish to Chemical Treatment
- Prepare caffeine exposure solutions by dissolving caffeine in reconstituted hard water. Perform appropriate serial dilutions by diluting higher caffeine treatments with hard water to produce lower caffeine treatment levels.
NOTE: Table 1 summarizes each of the treatment levels used in this experiment.
| Zebrafish | Fathead Minnow | ||||
| Treatment | Nominal Caffeine Concentration (mg/L) | Measured Caffeine Concentration (mg/L) | Treatment | Nominal Caffeine Concentration (mg/L) | Measured Caffeine Concentration (mg/L) |
| Control | 0 | <LOD | Control | 0 | <LOD |
| 75th Centile* | 0.001 | 0.001 | 75th Centile* | 0.001 | 0.001 |
| 95th Centile* | 0.039 | 0.013 | 95th Centile* | 0.039 | 0.009 |
| 99th Centile* | 0.412 | 0.361 | 99th Centile* | 0.412 | 0.310 |
| THV | 4.07 | 3.81 | THV | 4.07 | 4.12 |
| 10% LC50 | 48.46 | 46.66 | 10% LC50 | 14.1 | 14.7 |
| 40% LC50 | 193.82 | 186.67 | 40% LC50 | 56.38 | 53.91 |
Table 1: Experimental caffeine treatments for zebrafish and fathead minnow experiments. Nominal and measured values of caffeine for each treatment are given. *The caffeine treatments used in this study represent the upper centiles of exposure distributions based on measured environmental values of caffeine16. THV: Therapeutic Hazard Value. LOD: Limit of Detection
- Pour the prepared solution in individual exposure champers. Use 100 mL glass beakers filled with 20 mL of exposure solution for zebrafish exposure chambers and 500 mL beakers with 200 mL of exposure solution for fathead minnow exposure chambers.
- Using a transfer pipette, place 10 zebrafish embryos aged 4–6 h post fertilization (hpf) in each of four replicate exposure chambers per treatment.
- Place 10 fathead minnow larvae aged within 24 h of hatching in each of three replicate exposure chambers per treatment. To accommodate the larger size of fathead minnow larvae, cut the tip of the transfer pipette off prior to transfer.
- Maintain zebrafish experiments at a 16:8 h light:dark photoperiod and a constant temperature of 28 ± 1 °C. Use the same photoperiod regime for fathead minnow studies, but at a temperature of 25 ± 1 °C.
- After 96 h of chemical exposure, load individual fish in separate wells of 48 (for zebrafish) and 24 (for fathead minnow) well plates.
- To ensure that each well contains an equal volume of solution, transfer zebrafish larvae to 48 well plates using a 5,000 µL autopipette for a 1,000 µL volume per well. Use the autopipette to withdraw and transfer both the zebrafish larvae and exposure solution simultaneously.
- Due to their larger size, transfer fathead minnow larvae using a transfer pipette with the tip cut off. Prior to transferring fathead minnow larvae to individual wells, fill each well to 2,000 µL using an autopipette. When transferring individual fathead larvae to wells, place the tip of the transfer pipette in the well solution and allow the fish to swim from the pipette tip into the well.
2. Calibration of Video-tracking Parameters
- Prior to behavioral measures, set observation and calibration parameters in the video track software (see Table of Materials).
- Place a well plate in the recording chamber with at least 1 larval fish in an individual well. Use the plate and associated fish as representations to set calibration parameters.
- In the video track software, click "File | Generate Protocol", which will open a “protocol creation wizard” dialogue box. In the "Location Count" field, enter the number of individuals wells of the well plate and then click "OK".
- At the top of the screen, click "View | Full Screen", which will prompt the system to display an overhead camera view of the well plate.
- Click the "Draw Areas" icon, which appears as three multicolored shapes. To the right of the well plate viewing area, select the circle icon in the field labeled "Areas".
- Use the cursor to delineate the circular video tracking area in the top left well of the well plate. Select "Top-Right Mark" and then outline the viewing area of the top right well. Then, select "Bottom Mark" to outline the bottom right well.
NOTE: After drawing the circular outline, its position will likely need to be adjusted. To adjust the position of the outline, click “select” and then use the cursor to move the outlined area. Also, outlines can be replicated by clicking "Copy" and then "Paste". - After the top left, top right, and bottom right well tracking areas have been defined, click "Build" to prompt the software to automatically delineate the viewing areas of the remaining wells.
- In the area labeled "Calibration", click "Draw Scale". Use the cursor to draw a horizontal line across the plate. Once the line is drawn, a dialogue box labeled “Calibration measurement” will appear. Enter the well plate length and click "OK".
- Exit the drawing manager by clicking the "Draw Areas" icon.
- Click the "Tiles" icon. Using the cursor, highlight all the boxes that appear on the viewing screen so that each box is green.
NOTE: The Tiles icon appears as a group of six individual small squares - "Click View| Full Screen". To the right of the plate viewing area, click "Bkg" in the box labeled "Detection Threshold". Use the threshold adjustment bar to set the pixel detection threshold. Once, the appropriate pixel detection threshold is selected, click "Apply to Group".
NOTE: This protocol sets the detection threshold at 13 in black mode for zebrafish observations and at 110 in transparent mode for fathead minnow observations. - In the box labeled "Movement Threshold", enter the desired movement speed tracking parameters. Once speed parameters are set, click "Apply to Group".
NOTE: This protocol sets small/large movements at 20 mm/s and inactive/small movements at 5 mm/s. These selections program the software to track larval fish movement at three different speed levels: inactive (freezing) = <5 mm/s, small (cruising) = 5–20 mm/s, and large (bursting) = >20 mm/s. - Click "Parameters | Protocol Parameters" from the drop-down menu. In the dialogue box, select the "Time" tab. Enter the observation time and the integration time. After parameters are entered click "Ok".
- To set the light/dark photoperiod times and light intensity for each photoperiod open the light driver settings dialogue box by selecting "Light Driving" from the "Parameters" drop down menu.
NOTE: See protocol video for setting multiple light-dark photoperiods. - After the video tracking parameters have been set, save the observation protocol.
NOTE: This protocol observes fish behavior over a 50 min period that includes a 10 min acclimation phase followed by 4 altering light/dark phases consisting of two 10 min light periods and two 10 min dark periods. The integration time is set to measure behavior for each minute of the 50 min behavioral trial.
3. Observation of Larval Fish Locomotor and Photomotor Behavior
- Place the well plate containing experimental fish in the behavioral recording chamber.
- In the video tracking software, open the tracking protocol developed in step 3.
- In the video tracking viewer, check to make sure that all larvae are visible on the computer screen, that only one individual larva is present in each well, and that individual wells are aligned within the observation areas that were defined in steps 2.1.5 and 2.1.6.
- Click on "Experiment | Execute".
NOTE: The system will prompt the user to provide a name and location to save the observation data. - Once the name and save location of the observation data have been specified, click on the "Several Live Images" icon to highlight all the pre-defined viewing areas
NOTE: This icon is located at the top of the computer screen and appears as a box divided into four smaller squares. Clicking on this icon will highlight all the pre-defined viewing areas. - Close the panel of the recording chamber and click "Background | Start" on the computer monitor.
4. Analyzing Behavioral Data
- To retrieve larval fish activity data, open the spreadsheet, which is automatically compiled by the tracking software and is in the folder specified by the user before initiating behavioral trials (Step 3.4).
- Refer to Figures 1A and 1B for representative measurements of naive locomotor activity of unexposed zebrafish and fathead minnow larvae, respectively. Refer to Figures 1C and 1D for PMR calculations, which effectively examine the magnitude of movement difference between light to dark or dark to light transitions.
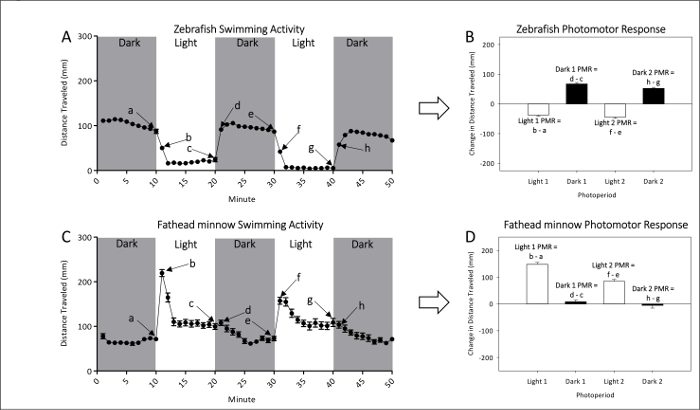
Figure 1: Example of baseline activity of unexposed zebrafish (A and B) and fathead minnow (C and D). The mean (± SEM) distance swam for zebrafish (A) and fathead minnow (C) is given by dots each representing one-minute intervals of activity. Two dark and two light periods of photomotor responses are measured. The last (a, c, e, and g) and first (b, d, f, and h) minute of each photoperiod are used to calculate PMRs. Photomotor responses of zebrafish (B) and fathead minnow (D) are measured as the change in mean (±SEM) distance traveled between the last minute of an initial photoperiod and the first minute of the following period. Please click here to view a larger version of this figure.
Caffeine treatment levels did not appreciably vary during the 96 h experiments with zebrafish and fathead minnows. For example, Table 1 presents analytically verified concentrations of each treatment level. This protocol verified water samples for caffeine treatment levels by isotope-dilution liquid chromatography tandem mass spectrometry (LC-MS/MS) generally following previously reported methods28. The formation of paraxanthine, the primary metabolite of caffeine, was also quantified. A description of these analytical procedures is provided in the supplemental analytical information. Because of the similarities between nominal and analytical verification of treatments, nominal treatment levels are presented throughout the remainder of this manuscript. Caffeine significantly altered zebrafish and fathead minnow behaviors. However, zebrafish locomotor responses were consistently more sensitive to caffeine than fathead minnows. The most sensitive behavioral endpoints for zebrafish and fathead minnow larvae were affected by caffeine at a concentration of 0.039 mg/L. Table 2 summarizes lowest observed effect concentrations (LOECs) and no observed effect concentrations (NOECs) for each behavioral endpoint in both fish models.
| Zebrafish | Fathead minnow | ||||
| Endpoint | LOEC (mg/L) | NOEC (mg/L) | Endpoint | LOEC (mg/L) | NOEC (mg/L) |
| Total Distance Dark | 0.412 | 0.039 | Total Distance Dark | − | 56.38 |
| Total Distance Light | 48.46 | 4.07 | Total Distance Light | − | 56.38 |
| Total Counts Dark | 0.412 | 0.039 | Total Counts Dark | − | 56.38 |
| Total Counts Light | 48.46 | 4.07 | Total Counts Light | − | 56.38 |
| Bursting Distance Dark | − | 193.82 | Bursting Distance Dark | − | 56.38 |
| Bursting Distance Light | 193.82 | 48.46 | Bursting Distance Light | − | 56.38 |
| Bursting Counts Dark | 193.82 | 48.46 | Bursting Counts Dark | − | 56.38 |
| Bursting Counts Light | 193.82 | 48.46 | Bursting Counts Light | − | 56.38 |
| Bursting Duration Dark | 193.82 | 48.46 | Bursting Duration Dark | − | 56.38 |
| Bursting Duration Light | − | 193.82 | Bursting Duration Light | − | 56.38 |
| Cruising Distance Dark | 0.412 | 0.039 | Cruising Distance Dark | − | 56.38 |
| Cruising Distance Light | 48.46 | 4.07 | Cruising Distance Light | − | 56.38 |
| Cruising Counts Dark | 0.412 | 0.039 | Cruising Counts Dark | − | 56.38 |
| Cruising Counts Light | 48.46 | 4.07 | Cruising Counts Light | − | 56.38 |
| Cruising Duration Dark | 0.412 | 0.039 | Cruising Duration Dark | − | 56.38 |
| Cruising Duration Light | 48.46 | 4.07 | Cruising Duration Light | − | 56.38 |
| Freezing Distance Dark | 0.412 | 0.039 | Freezing Distance Dark | 0.039 | 0.001 |
| Freezing Distance Light | 0.039 | 0.001 | Freezing Distance Light | − | 56.38 |
| Freezing Counts Dark | 0.412 | 0.039 | Freezing Counts Dark | − | 56.38 |
| Freezing Counts Light | 48.46 | 4.07 | Freezing Counts Light | − | 56.38 |
| Freezing Duration Dark | − | 193.82 | Freezing Duration Dark | 56.38 | 14.10 |
| Freezing Duration Light | 48.46 | 4.07 | Freezing Duration Light | − | 56.38 |
| Dark 1 PMR | 48.46 | 4.07 | Dark 1 PMR | 0.039 | 0.001 |
| Light 1 PMR | 48.46 | 4.07 | Light 1 PMR | − | 56.38 |
| Dark 2 PMR | 48.46 | 4.07 | Dark 2 PMR | − | 56.38 |
| Light 2 PMR | 48.46 | 4.07 | Light 2 PMR | − | 56.38 |
| Bursting Dark 1 PMR | − | 193.82 | Bursting Dark 1 PMR | − | 56.38 |
| Bursting Light 1 PMR | − | 193.82 | Bursting Light 1 PMR | − | 56.38 |
| Bursting Dark 2 PMR | 193.82 | 48.46 | Bursting Dark 2 PMR | − | 56.38 |
| Bursting Light 2 PMR | − | 193.82 | Bursting Light 2 PMR | − | 56.38 |
| Cruising Dark 1 PMR | 48.46 | 4.07 | Cruising Dark 1 PMR | − | 56.38 |
| Cruising Light 1 PMR | 48.46 | 4.07 | Cruising Light 1 PMR | − | 56.38 |
| Cruising Dark 2 PMR | 48.46 | 4.07 | Cruising Dark 2 PMR | − | 56.38 |
| Cruising Light 2 PMR | 193.82 | 48.46 | Cruising Light 2 PMR | 56.38 | 14.10 |
| Freezing Dark 1 PMR | 48.46 | 4.07 | Freezing Dark 1 PMR | − | 56.38 |
| Freezing Light 1 PMR | 193.82 | 48.46 | Freezing Light 1 PMR | − | 56.38 |
| Freezing Dark 2 PMR | 48.46 | 4.07 | Freezing Dark 2 PMR | − | 56.38 |
| Freezing Light 2 PMR | 193.82 | 48.46 | Freezing Light 2 PMR | − | 56.38 |
Table 2: Zebrafish and fathead minnow behavioral NOECs and LOECs for Caffeine. No Observed Effect Concentration (NOEC) and Lowest Observed Effect Concentration (LOEC) (mg/L) values for each of the light/dark swimming activity endpoints and photomotor responses for zebrafish and fathead minnows exposed to caffeine. Dashes indicate that no effects were observed at a particular endpoint across all treatment levels.
Figure 2 presents total locomotor activity and PMRs of zebrafish and fathead minnow following 96 h exposure to caffeine. Fathead minnow larvae PMRs were altered by caffeine at lower treatment levels (0.038 mg/L) than zebrafish, but a markedly larger number of photomotor endpoints were affected in zebrafish. The highest treatment level of caffeine (193.82 mg/L) altered PMR in zebrafish, in which these responses were exactly opposite from controls. At this elevated treatment level, however, PMRs decreased in dark and increased in light conditions.
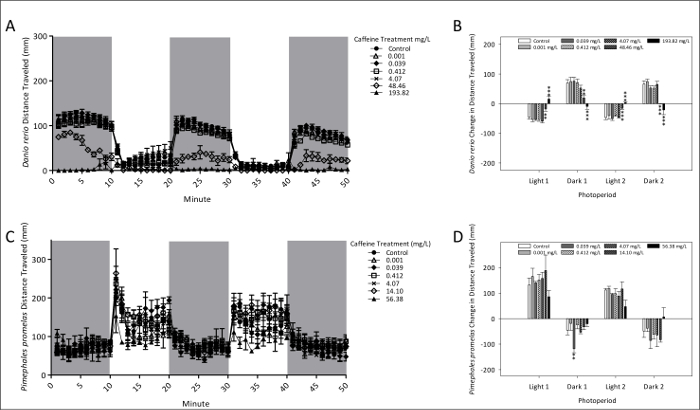
Figure 2: Swimming activity and photomotor responses of zebrafish (A and B) and fathead minnow (C and D) after 96 h exposure to caffeine. The mean (± SEM) distance swam for zebrafish (A) and fathead minnow (C) is given by dots each representing 1-min intervals of activity. Photomotor responses of zebrafish (B) and fathead minnow (D) are measured as the change in mean (± SE) total distance traveled between the last minutes of an initial photoperiod and the first minute of the following period. Two dark and two light period photomotor responses were measured. A total of 24 (4 replicates each of 6 larvae) zebrafish and 12 (3 replicates each of 4 larvae) fathead minnows were used for behavioral observation. *p <0.10; ** p <0.05; *** p <0.01. Please click here to view a larger version of this figure.
In addition to measuring larval PMRs, light and dark locomotor activity was analyzed across three speed thresholds for distance moved, number of movements, and duration of movements. This data is used to develop behavioral response profiles for caffeine (Figure 3, Supplemental Figure 1). In both of the fish models, caffeine inhibited activity at all significantly affected locomotor endpoints. Both fish models demonstrated increased activity at the bursting speed thresholds following exposure to caffeine, though not significantly. Similar to the results of the PMR observations, caffeine effected a greater number of zebrafish locomotor endpoints. In fact, caffeine significantly altered several locomotor responses under dark conditions at environmentally realistic levels below the THV. However, fathead minnow locomotor activity was not significantly affected under light conditions by any treatment level.

Figure 3: Response profiles of larval zebrafish and fathead minnows after 96 h exposure to caffeine. Mean zebrafish dark (A) and light (B) swimming activity compared to mean fathead minnow dark (C) and light (D) activity after 96 h exposure to caffeine. Plotted data represents activity over two 10 min dark photoperiods and two 10 min light photoperiods for each fish model. Data is normalized to control, which is represented at the 0 axis in each figure. Behavioral parameters include distance swam, number of movements (counts), and duration of each movement across 3 speed levels, bursting (>20 mm/s), cruising (5–20 mm/s), and freezing (<5 mm/s). In addition to movement patterns at each of the speed thresholds, total distance swam, and total number of movements is represented. ↑ represents a significant increase in activity in comparison to control and ↓ indicates a significant decrease in activity in comparison to control. A total of 24 (4 replicates each of 6 larvae) zebrafish and 12 (3 replicates each of 4 larvae) fathead minnow where used in behavioral observations for each group. *p <0.10; ** p <0.05; *** p <0.01. Please click here to view a larger version of this figure.
Supplemental Figure 1: Photomotor responses of zebrafish (A and B) and fathead minnow (C and D) across three speed thresholds. Zebrafish (A, B, and C) and fathead minnow larvae (D, E, and F) photomotor responses across three speed thresholds (Freezing: 20 mm/s) after 96hr exposure to caffeine. Photomotor responses of zebrafish and fathead minnow are measured as the change in mean (±SE) total distance traveled between the last minutes of an initial photoperiod and the first minute of the following period. Two dark and two light period photomotor responses were measured. A total of 24 (4 replicates each of 6 larvae) zebrafish and 12 (3 replicates of 4 larvae) fathead minnows were used for behavioral observation. *p <0.01 Please click here to download this file.
