The Caenorhabditis Sieve consists of 2 screw caps, securing an area of woven nylon monofilament mesh smaller than the body diameter of the desired developmental age, used to extract live populations of organisms using a simple washing technique. It attaches to standard conical tubes and uses the mesh screen to mechanically sort animals by body diameter, leaving the desired animals in the tube ready for further maintenance and experimentation (e.g., transfer or genetic harvest). A gentle manual washing with the Caenorhabditis Sieve is quick, approximately 5 min per 60–100 mm plate, and the organisms are easily recovered from the mesh.
Percentage yield of animals following Caenorhabditis Sieve use
To establish the percentage yield of the Caenorhabditis Sieve, devices were tested with both 20 μm and 50 μm pore-size meshes on adult animals. A mean yield of a >90% successful animal transfer was achieved for both mesh sizes tested (Table 1).
Monofilament mesh with different size gaps may be used to separate animals of different life stages. Mesh with 20 μm gaps is appropriate for washing away developing embryos and larval stages smaller than the fourth larval stage, retaining the latter (L4; with an average body diameter of 32 μm) and any animals in later life stages, while mesh with 50 μm gaps will allow all other life stages aside from adults (with an average body diameter of 70 μm) to be washed away (Table 2).
Caenorhabditis Sieve use does not impact healthspan metrics
Motility: In C. elegans the normal sinusoidal movement (i.e., motility) declines with age18 and is a marker of overall health. To determine if the Caenorhabditis Sieve influenced motility, motility scores were compared for pick, FUDR, and Caenorhabditis Sieve treatment groups on days 2, 4, 6, and 8 of adulthood. All the animals across every group (n = 10/group) exhibited normal and spontaneous movement patterns (Class A) at multiple ages throughout adulthood (days 2, 4, 6, and 8 of adulthood; p > 0.05 for multiple comparisons on all days, Figure 3).
Pharyngeal pump rate: The ability of C. elegans' pharyngeal muscles to pump declines with age and is another biomarker of healthspan23. To determine if the Caenorhabditis Sieve influenced the animals' pharyngeal pump rate, pick, FUDR, and Caenorhabditis Sieve treatment groups were compared on days 2, 4, 6, and 8 of adulthood (n = 8 to 10 per group). There was a significant difference between the animals that underwent the picking and FUDR methods on days 6 (p <0.001) and 8 (p <0.001). Also, there was a significant difference between the sieve and FUDR groups on days 6 (p <0.001) and day 8 of adulthood (p <0.001). However, there was no statistically significant difference between the pick and the Caenorhabditis Sieve groups for any day (p >0.05, Figure 4), indicating that the sieve does not impact this measure of healthspan.
Gentle touch response: Response to mechanical stimulus is a physiological marker to assess aging or general health24,25; thus, the impact of different transfer methods on both the anterior and the posterior gentle touch responses were tested. There was no statistically significant difference between the pick, FUDR, and Caenorhabditis Sieve treatment groups (n = 8/group), either anteriorly or posteriorly, for any day of testing (p >0.4 for all comparisons; Figure 5A and 5B).
Fecundity: To establish whether or not the Caenorhabditis Sieve influenced the amount of viable progeny produced by C. elegans, the individual offspring produced in a 24 h period during day 3 of adulthood was counted and compared (n = 20 to 22 per group). The use of the Caenorhabditis Sieve did not significantly impact the number of progeny produced when compared to a pick treatment group (p = 0.61, Figure 6).
Molecular reporter assays
DAF-16 nuclear translocation: In C. elegans, the activation of the transcription factor DAF-16 is associated with increased stress resistance26. The nuclear localization of DAF-16 was examined in a transgenic nematode strain TJ356, which expresses DAF-16 fused to a green fluorescent protein (DAF-16::GFP)20. Under normal growth conditions, DAF-16::GFP is localized primarily in the cytosol, but under various stressors (e.g., heat stress), it is rapidly translocated into the nucleus20. To test the impact of sorting with the Caenorhabditis Sieve on DAF-16 translocation, DAF-16::GFP localizations were compared in age-matched day-5 adults in a positive control group (heat stress), a negative control group (manual transfer via pick), and a Caenorhabditis Sieve treatment groups (n = 10/group). The transfer with the Caenorhabditis Sieve did not affect the nuclear translocation of DAF-16::GFP, and showed a similar phenotype to the negative control animals (p >0.05, Figure 7).
hsp-16.2 reporter: Small heat shock proteins like HSP-16.2 are biomarkers of a stress response, and they are highly expressed during an exposure to heat shock or oxidative stress agents21,27. The TJ375 strain has a GFP reporter gene fused with an hsp-16.2 promoter that is not active under normal conditions21. However, after an exposure to a heat shock, HSP-16.2 protein expression is induced, and the animals display high levels of GFP expression21. To test the involvement of the Caenorhabditis Sieve on an HSP-16.2-mediated stress response, the fluorescence density in the pharynx region (n = 10 animals/group) of age-matched animals was compared in day-5 adults between a positive control group (heat stress), a negative control group (picking), and a Caenorhabditis Sieve treatment group. The transfer with the Caenorhabditis Sieve did not significantly induce the expression of HSP-16.2::GFP (hsp-16.2::gfp) when compared to the negative control (p >0.05, Figure 8).
sod-3 reporter: In C. elegans, the anti-oxidant gene that codes for superoxide dismutase 3 (SOD-3) is up-regulated during oxidative stress28. The C. elegans strain CF1553 expresses green fluorescent protein (GFP)-labeled SOD-3 promoter, whose expression is induced by oxidative stressors, such as paraquat5. To test the involvement of the Caenorhabditis Sieve sorting on the antioxidant response in C. elegans, the fluorescence density in the head region of age-matched day-5 adults was compared between a positive control population (100 μM paraquat treatment), a negative control population (manual picking), and a Caenorhabditis Sieve-transferred population. The transfer with the Caenorhabditis Sieve did not significantly induce the expression of sod-3::gfp when compared to the negative control (p >0.05, Figure 9).

Figure 1: Caenorhabditis Sieve construction. The progression of the 'do-it-yourself' manufacture of the tool is shown. These panels show (A) two 50 mL conical tube caps (B) whose centers have been removed, (C) cut edges smoothed, and (D – F) who are fitted with monofilament mesh corresponding to the desired life stage of C. elegans (see Protocol for details). (G) The completed sieve is also shown. Please click here to view a larger version of this figure.

Figure 2: Step-by-step image representation of Caenorhabditis Sieve use. (A) The sieve is pre-wet with a drop of M9 solution, and (B) fit on top of a 50 mL conical tube. (C) 1 mL of M9 solution with worms is pipetted on the topside of the sieve, (D) a 50 mL conical tube is placed on top of the sieve with the worms facing the inside of the tube, and (E) the sieve with a newly attached upper tube is quickly flipped over. (G) The sieve is rinsed with M9 carrying the desired animals into the new 50 mL tube and the worms are allowed to settle by gravity to the bottom of the tube. (H) The worms are pipetted and placed as droplets on a fresh NGM plate. Please click here to view a larger version of this figure.
Figure 3: Caenorhabditis Sieve did not impact motility throughout lifespan. This figure shows the motility class distribution of animals at days 2, 4, 6, and 8 of adulthood for pick, FUDR, and Caenorhabditis Sieve treatment groups. Class A animals moved normally and spontaneously, class B animals moved abnormally and may have required prodding, and class C animals were unable to move. There was no difference between Caenorhabditis Sieve, pick and FUDR groups (p >0.05). Three replicates (n = 10 animals per treatment plate) were conducted and analyzed with Ordinal logistic model. class B animals. Please click here to view a larger version of this figure.
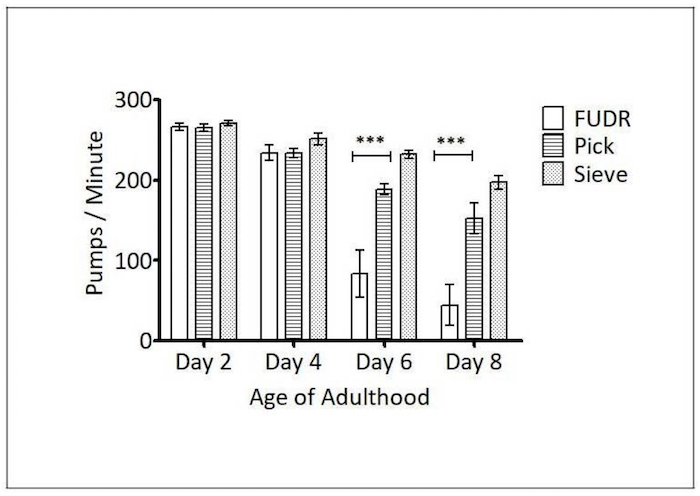
Figure 4: Caenorhabditis Sieve use did not impact pharyngeal pumping throughout the lifespan. This figure shows the pharyngeal pump rates of pick, FUDR, and Caenorhabditis Sieve treatment groups, compared on days 2, 4, 6, and 8 of adulthood. The asterisks denote a significance between the pick, sieve, and FUDR treatment groups for the days specified (*** p <0.05). There was no difference between the Caenorhabditis Sieve and the pick group (p > 0.05). Two replicates (N = 10 animals per treatment plate) were conducted and analyzed with a one-way ANOVA and a Bonferroni post-test. The bars represent the mean ± the standard error of the mean. Please click here to view a larger version of this figure.
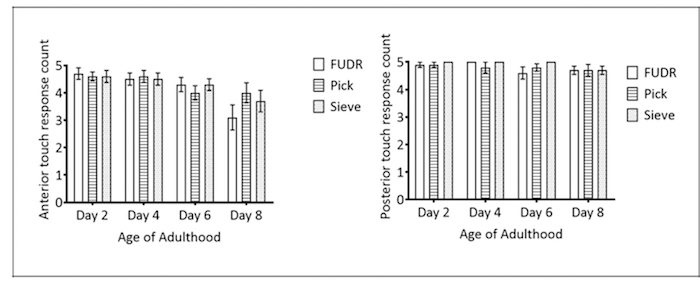
Figure 5: Caenorhabditis Sieve use did not impact anterior touch response throughout lifespan. These panels show (A) the anterior and (B) the posterior touch response count of pick, FUDR, and Caenorhabditis Sieve treatment groups compared on days 2, 4, 6, and 8 of adulthood. Two replicates were conducted with N = 10 for each treatment group and compared with a one-way ANOVA and a Bonferroni post-test (p = 0.4 and p = 0.9 for anterior and posterior, respectively). The bars represent the mean ± the standard error of the mean. Please click here to view a larger version of this figure.
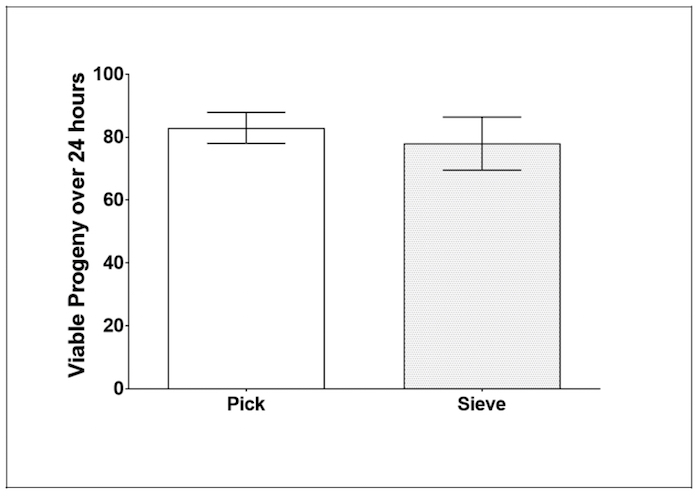
Figure 6: Caenorhabditis Sieve did not impact the amount of viable progeny on day 3 of adulthood. This figure shows the viable progeny of day-3 adults after a 24 h egg-laying period, spanning day 3 of adulthood for parent animals. The bars represent the mean ± the standard error of the mean. N = 20 – 22 animals from at least two separate biological replicates per treatment group. The treatment groups are compared with a t-test, (p >0.05). Please click here to view a larger version of this figure.
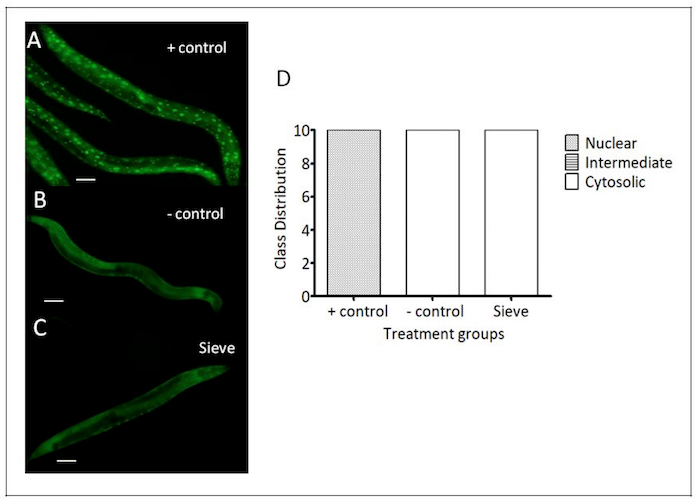
Figure 7: Caenorhabditis Sieve did not affect nuclear translocation of DAF-16::GFP. These panels show representative images of the DAF-16 translocation of (A) a heat shock group (the positive control), (B) a pick group (the negative control), and (C) a Caenorhabditis Sieve treatment group. (D) The animals in the positive control group displayed an activation of the DAF-16 nuclear translocation. The Caenorhabditis Sieve did not induce a nuclear translocation and displayed cytosolic fusion protein similar to the animals in the negative control group. N = 10 animals per treatment group from at least three separate biological replicates. The treatment groups were compared with a one-way ANOVA and a Tukey's post hoc test. Scale bar = 100 μm. Please click here to view a larger version of this figure.
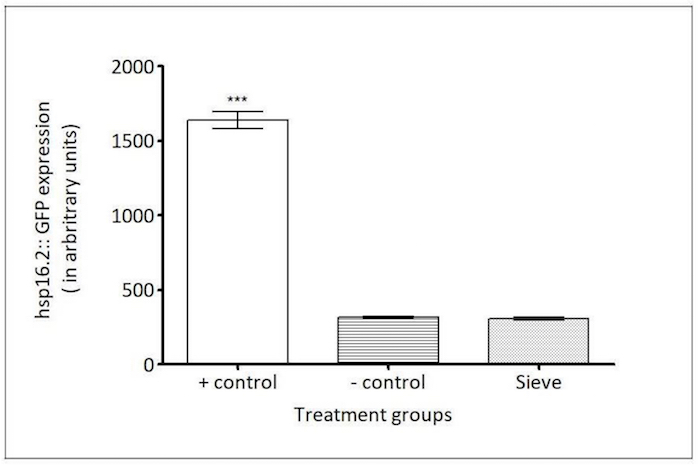
Figure 8: Caenorhabditis Sieve did not affect the expression of hsp16.2::gfp. The HSP-16.2 expressions (in arbitrary fluorescence units) of a heat shock group (the positive control), a pick group (the negative control), and a Caenorhabditis Sieve treatment group are compared. The asterisks denote a high expression of hsp16.2::gfp for the positive control group which was significantly different from the other treatment groups (*** p <0.05). The Caenorhabditis Sieve did not affect the hsp16.2::gfp expression and displayed fluorescence intensities similar to animals in the negative control group (p >0.05). Three replicates were conducted with N = 10 for each treatment group and compared with a one-way ANOVA and a Tukey's post hoc test. Please click here to view a larger version of this figure.
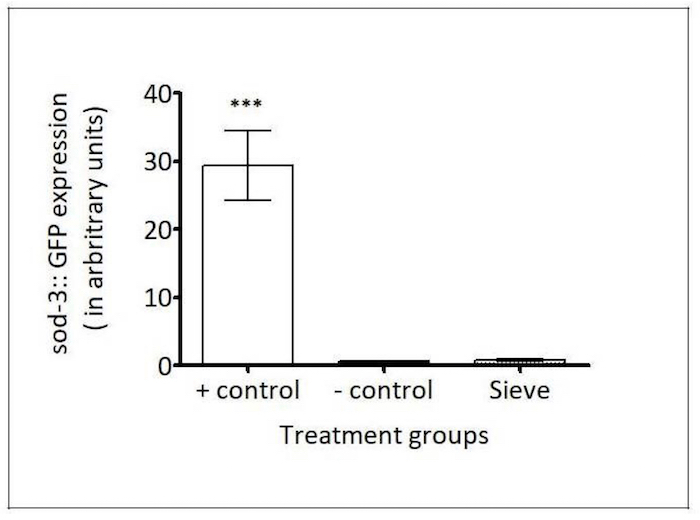
Figure 9: Caenorhabditis Sieve did not affect the expression of sod-3::gfp. The SOD-3 expressions (in arbitrary fluorescence units) of a 100 μM paraquat group (the positive control), a pick group (the negative control), and a Caenorhabditis Sieve treatment group are shown. The asterisks denote a high expression of sod-3::gfp for the positive control group which was significantly different from the other groups (*** p <0.05). The Caenorhabditis Sieve did not affect the sod-3::gfp expression and displayed fluorescence intensities similar to the animals in the negative control group (p >0.05). Three replicates were conducted with N = 10 for each treatment group and compared with a one-way ANOVA and a Tukey's post hoc test. Please click here to view a larger version of this figure.
| Mesh size | N = 50 | N = 100 |
| 20 μm | 95.33% | |
| 50 μm | 99.00% | 93.33% |
Table 1: Percentage yield of mesh sizes. This table shows the results of a 20 μm device tested with N = 50 adults for 3 replicates and a 50 μm device tested with N = 50 adults for 2 replicates and with N = 100 for 3 replicates.
| Mesh Size | Developmental Stage | Body Diameter |
| 20 μm | Larval Stage 4 | ~35 μm |
| 50 μm | Day 1 Adult | ~70 μm |
Table 2: Mean body diameter. This table displays the mean body diameter acquired by averaging 3 measurements equally spread across each worm for 15 animals at each measured life stage.
