All zebrafish experiments have been approved by the Animal Experiment Board in Finland (ESAVI/8245/04.10.07/2015). Methods are performed according to the act (497/2013) and the government decree (564/2013) on the protection of animals used for scientific or educational purposes in Finland.
1. Culturing of Mycobacterium marinum
NOTE: Since Mycobacterium marinum is a pathogen capable of causing superficial infections in humans, find out the local guidelines for personal safety and biohazard waste disposal before starting to work with this bacterium.
- Culture M. marinum on a 7H10 plate supplemented with 10% OADC (oleic acid, albumin, dextrose, catalase) enrichment and 0.5% v/v of glycerol at 29 °C for at least 5 days. Maintain M. marinum cultures by taking fresh bacteria from the freezer every two weeks and transferring onto a new plate every other week.
NOTE: M. marinum is a natural fish pathogen infecting aquatic species and it is important to take precautions not to contaminate zebrafish stocks with the bacteria. Infected zebrafish and items contaminated with bacteria have to be kept separate from the breeding facilities. - Use a sterile 1-µL inoculation loop to aseptically transfer a loopful of M. marinum bacterial mass into a cell culture flask containing 10 mL of 7H9 medium with 10% ADC (albumin, dextrose, catalase) enrichment, 0.2% v/v polysorbate 80 and 0.2% v/v of glycerol. Culture for 3–4 days to approximately an OD600 (optical density) of 0.7 at 29 °C in the dark without shaking. Leave the cap loose, or use a filter cap, to allow for the sufficient exchange of gases.
NOTE: Polysorbate 80 is added to the medium to prevent the aggregation of bacteria. In addition, exposure to light leads to phenotypic changes in bacterial colonies (e.g., the color changes from white to yellow). To avoid this, keep the cultures in the dark. - Measure the OD600value with a spectrophotometer. Dilute the liquid culture to an OD600 of 0.07–0.09 and continue culturing for 2 days at 29 °C in the dark without shaking. Leave the cap loose.
NOTE: During these two days, the bacterial suspension will reach an OD600of approximately 0.5 corresponding to an early log-phase.
2. Preparation of Bacterial Solution for Infecting Adult Zebrafish
- Transfer the bacterial suspension into a big sterile cuvette or a 15 mL tube and place it in the dark at room temperature for 15 min to allow the biggest clumps to settle.
- Transfer the top 5–7 mL of the suspension into a clean tube or a cuvette and measure the OD600. Use this top phase of the suspension for the infections.
- Collect 1 mL of M. marinum culture into a fresh tube and centrifuge for 3 min at 10,000 x g. Remove the supernatant and resuspend the pellet in 1 mL of sterile 1x PBS.
- Dilute to reach the desired bacterial concentration by using sterile 1x PBS with 0.3 mg/mL phenol red as a tracer. Divide the diluted suspension into three aliquots.
NOTE: Use a predetermined OD600 vs. CFU (colony-forming units)/µL curve to estimate the dilution required to get the wanted number of bacteria in the 5 µL injection volume34. The correlation between OD600 and concentration of the bacterial suspension needs to be validated before starting the actual infection experiments. Reserve two weeks for collecting this validation data. - Using a 1 mL syringe, slowly pull the suspension through a 27 G needle 3x. For each aliquot, perform this step just before use.
NOTE: Do not use the same bacterial solution for more than 2 h.
3. Experimental M. marinum Infection with Intraperitoneal Injection
- Pipette a 5 µL droplet of the diluted bacterial solution onto a piece of parafilm film and pull the droplet into a 30 G insulin needle.
- Use 5–8 month-old wild-type fish and rag1−/−hu1999 mutant fish for the experiment. Anesthetize adult zebrafish in the tank water with 0.02% 3-aminobenzoic acid ethyl ester (pH 7.0). Position the fish ventral side up into a slit on a moist foamed plastic.
NOTE: Rag-/- mutant fish are not able to undergo somatic recombination and produce functional T and B cells. - Inject the needle between the pelvic fins at an approximately 45° angle. Keep the needle opening upwards to observe that the entire opening is inside the abdominal cavity. Slowly inject the bacterial suspension and carefully remove the needle.
NOTE: In case the red tracer is leaking out of the fish upon injection, exclude the fish from the experiment. - Immediately after injection, transfer the fish into a recovery tank with fresh tank water.
- Take samples of the bacterial suspension on 7H10 plates every 15 min from the bacterial aliquot in use and incubate the bacteria at 29 °C for 5 days and verify the infection dose by counting the colonies on the plates.
- Check the well-being of the fish regularly and euthanize any fish with symptoms of the infection with over 0.02% concentration of 3-aminobenzoic acid ethyl ester (pH 7.0).
NOTE: Approximately, 7% of the adult zebrafish infected with 34 ± 15 CFU and 30% of zebrafish infected with 2029 ± 709 CFU will have had symptoms by 8 weeks19. The symptoms may include abnormal swimming, lack of response to touch, gasping, edema or observable wasting. - Maintain the zebrafish according to the common standards35.
4. Collection of Internal Organs
- Euthanize the zebrafish with an overdose of 3-aminobenzoic acid ethyl ester (over 0.02% concentration, pH 7.0) in the tank water.
- Insert one pin posterior to the branchiostegal rays and another through the tail to tack the fish onto the platform.
- Open the whole abdominal cavity with a scalpel and collect the internal organs by using a small spoon and sharp-ended tweezers. Start from the heart and work along the spine towards the tail to detach all internal organs in one block.
NOTE: Be sure to collect all kidney tissue by scraping along the spine with the spoon. - Finally, use tweezers to detach the gut next to the cloaca and transfer the organs into a 1.5 mL homogenization tube with six 2.8 mm ceramic beads. Immediately place on dry-ice to freeze the sample. The sample can be stored at -80 °C until homogenized.
- Rinse the instruments with 70% ethanol between individuals.
5. Homogenization and RNA extraction from an organ block.
NOTE: The method is modified from the Stanford University protocols36.
- Add guanidine thiocyanate-phenol solution used for nucleic acid extraction (Table of Materials) on the top of the sample to a total volume of 1,500 µL. Ensure that the sample covers a maximum of 10% of the total volume.
CAUTION: The expected volume of the harvested tissue is 100 µL. Guanidine thiocyanate-phenol solution contains toxic and irritating compounds and requires protective clothing, nitrile gloves and working in a fume hood. Do not combine with bleach as this will cause formation of toxic gases. Read the material safety data sheet (MSDS) before use. - Homogenize samples using a bead-beating homogenizer 3 times for 40 s at 3,200 rpm. Cool on ice for 30 s between the cycles. Sonicate the homogenized samples in a water bath for 9 min.
- Centrifuge the samples at 12,000 x g for 10 min at 4 °C and move 1,000 µL of the cleared homogenate into a fresh microcentrifuge tube.
- Add 200 µL of chloroform, immediately mix by vortexing for 15 s and incubate for 2 min at room temperature.
CAUTION: Chloroform is a toxic and irritant compound if inhaled, swallowed or contacted with skin or eyes. Use necessary safety equipment for personal protection and work in a fume hood. Read the material safety data sheet (MSDS) before use. - Centrifuge at 12,000 x g for 15 min at 4 °C to separate the aqueous and organic phases.
- Carefully, transfer 500 µL of the top phase to a fresh tube to avoid contaminating the RNA. Remove and discard the rest of the aqueous phase (~100 µL) and store the interphase and organic phase at 4 °C for DNA extraction.
NOTE: The top phase contains the RNA. - Add 500 µL of 2-propanol and immediately mix by vortexing for 15 s. Incubate for 10 min at room temperature to precipitate the RNA.
- Centrifuge at 12,000 x g for 10 min at 4 °C to pellet the RNA. Remove the supernatant by pipetting.
- Add 1 mL of 75% ethanol and vortex for 10 s.
NOTE: The protocol can be paused here, and the samples kept overnight at 4 °C. - Centrifuge at 7,500 x g for 5 min at 4 °C. Remove the supernatant by pipetting.
- Repeat the wash steps 5.9–5.10. Remove the supernatant carefully by pipetting and let the pellet air-dry in a fume hood.
- Dissolve the RNA pellet in 500 µL of nuclease-free water and keep the samples on ice. Measure the concentrations with a microvolume spectrophotometer or with equivalent equipment. Store the RNA at -80 °C.
6. Purification of Co-extracted Zebrafish and Mycobacterial DNA
- Prepare a back-extraction buffer (BEB) by dissolving 118.2 g of guanidine thiocyanate (final concentration 4 M), 3.68 g of sodium citrate (final concentration 50 mM) and 30.29 g of Tris free base (final concentration 1 M) in 120 mL of nuclease-free water (this may require stirring overnight). Add nuclease-free water to a final total volume of 250 mL and filter to sterilize the solution.
NOTE: This buffer can be stored at room temperature for up to 6 months. Do not combine BEB with bleach as they react to produce toxic gases such as hydrogen chloride and hydrogen cyanide. - Use the interphase and organic phase of the sample to extract mycobacterial DNA. Add 500 μL of BEB to each tube. Mix extensively for 10 min by inversion at room temperature.
- Centrifuge the tubes at 12,000 x g for 30 min at room temperature and carefully transfer 500 µL of the upper aqueous phase containing the DNA to a new tube.
- Add 400 μL of 2-propanol. Mix by inverting and incubate for 10 min at room temperature.
- Centrifuge the samples at 12,000 x g for 15 min at 4 °C. A pellet containing the DNA should be visible at this point. Carefully remove the supernatant by pipetting.
- Add 800 μL of 70% ethanol. Wash the pellet by inversion. Do not vortex the samples at this point, as genomic DNA breaks down easily.
- Centrifuge the samples at 12,000 x g for 15 min at 4°C and remove the supernatant by pipetting. Repeat the ethanol wash (steps 6.6 and 6.7).
- Remove the ethanol by carefully pipetting. Let the samples air-dry for 5–10 min. Dissolve the pellet in 200 μL of nuclease-free water.
- Measure DNA concentrations with a microvolume spectrophotometer or with equivalent equipment. DNA can be stored at 4 °C or at -20 °C for long-term storage.
7. Quantitative PCR for Measuring Mycobacterial Loads
- Prepare qPCR reaction mixes with no-ROX (carboxy-X-rhodamine) against the M. marinum internal transcribed spacer (ITS) between 16S-23S ITS according to the manufacturer's instructions with MMITS1 primers (Table 1). Pipette the reaction mix and sample dilutions as duplicates on a 96-well plate suitable for qPCR. Include a DNA standard dilution series of a known amount of bacteria in each run.
NOTE: The expected M. marinum load per fish can range from 0 CFU to 1,000,000 CFU at 4 wpi. The qPCR assay can be also performed with other qPCR kits but the annealing temperature for the primers has to be re-optimized. - Seal the plate with an optically transparent film and centrifuge the plate at 2,000 x g for 2 min at 4 °C.
- Run the qPCR program shown in the Table 2.
- Using the standard curve, calculate the number of bacteria in the entire fish sample.
8. DNase Treatment of the RNA Samples
- To remove any possible remaining traces of genomic DNA from the RNA, carry out DNase I treatment. Thaw the RNA samples on ice.
NOTE: Be sure to use only RNase-free equipment and solutions and wipe the working surface and pipettes with a decontamination reagent eliminating RNases (Table of Materials) before starting to work. Wear a long-sleeved lab coat and gloves to protect your samples. - Prepare 10 μL DNase I reaction mixes on ice according to the manufacturer's instructions. Mix 1 μL of DNase I, 1 μL of 10x DNase buffer and 8 μL of RNA sample including a maximum of 1 μg of RNA.
- Gently mix the reactions (no vortexing) and incubate for 30 min at 37 °C.
- Prior to heat-inactivation, add 1 µL of 50 mM EDTA to each 10 µL sample. If EDTA is not added, the RNA will undergo chemical degradation when heated.
- Incubate for 10 min at 65 °C to heat-inactivate DNase I. Continue directly to cDNA synthesis or store the DNase-treated RNA at -80 °C.
9. cDNA Synthesis
- Keep all reagents and samples on ice and prepare the reaction mixes according to the manufacturer's instructions. For a 5 μL reaction mix, include 1 μL of Reverse Transcription Master Mix, 3 μL of nuclease-free water and 1 μL of DNase treated RNA.
- Gently mix the reverse transcription reactions and briefly spin the tube, if needed.
- Place samples in a PCR machine and use the program shown in Table 3.
- Dilute the cDNA in nuclease-free water for qPCR to a maximum concentration of 2.5 ng/μL, if needed. cDNA can be stored at -20 °C.
10. Measuring Zebrafish Gene Expression by Quantitative PCR
- Prepare a qPCR master mix on ice according to the manufacturer's instructions and protect from light. Use the primers introduced in Table 1.
NOTE: To calculate the fold of induction for each gene, measure the expression also from a pooled baseline sample extracted from 6 healthy zebrafish. - Prepare replicates of each sample and pipette the reaction mixes onto a qPCR plate. Seal the plate with an optically transparent film and centrifuge the plate at 2,000 x g for 2 min at 4 °C before starting the run.
- Run the qPCR program shown in the Table 4 with the annealing temperature depending on the primer pair used (Table 1).
- Analyze the gene expression ratio compared to a house-keeping gene (loopern437) with the ΔCt method using the equation:
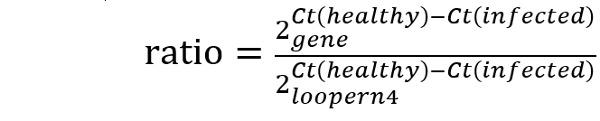
The natural fish pathogen Mycobacterium marinum infects the internal organs of the zebrafish and produces a systemic infection with histologically visible granulomas19. Adult zebrafish are infected with M. marinum by an intraperitoneal injection. The DNA and RNA are extracted, and the mycobacterial load is measured by quantitative polymerase chain reaction (qPCR) using DNA as the template. The outline of the method is shown in Figure 1.
The initial number of mycobacteria used for infecting the fish is a critical determinant for the outcome of infection. A high infection dose of M. marinum (~2,000 CFU) leads to a progressive disease in which the mycobacterial loads continue to increase until the average bacterial load reaches around five million bacteria (Figure 2A) ultimately killing the fish. A low dose (~20–90 CFU) of M. marinum leads to the development of a disease spectrum similar to that seen in human tuberculosis (Figure 2B). The bacterial load continues to increase until around 4–7 weeks (Figure 2A and Figure 3A), after which in the majority of the fish the disease reaches a steady-state. Figure 2B shows an example of the distribution of disease outcomes with a low dose infection: About 7% of the infected zebrafish were unable to restrict the bacterial growth. These individuals developed a primary progressive disease and they died within two months after the infection. Around 10% of the individuals cleared the mycobacterial infection by 4 weeks. The remaining 65% of the fish population developed a latent mycobacterial infection with steady bacterial burdens. However, between 8 and 32 weeks of infection, in 18%, the latent infection spontaneously reactivated leading to the progression of the disease.
By using rag-/- mutant fish, it is possible to study the role of adaptive immune responses in the adult fish. Rag-/- mutant fish cannot sufficiently limit the growth of mycobacteria leading to higher bacterial loads (Figure 3A) and increased morbidity (Figure 3B), clearly demonstrating the importance of adaptive immunity in controlling mycobacterial infection. Also, the importance of adaptive responses in evoking certain cytokine responses in mycobacterial infection can be studied in this model. Here, we show that the adaptive response is required for the efficient induction of interleukin 4 (IL4) (Figure 3C) but is dispensable for the induction of interferon-γ (IFNγ) at 4 wpi (Figure 3D). Interferon-γ is a cytokine driving the response against intracellular pathogens whereas interleukin 4 is a common mediator in the adaptive immune response against extracellular pathogens. The significantly higher expression levels of il4 in the wild-type group compared to rag-/- mutant fish refers to important adaptive humoral responses in the mycobacterial infection (Figure 3C).
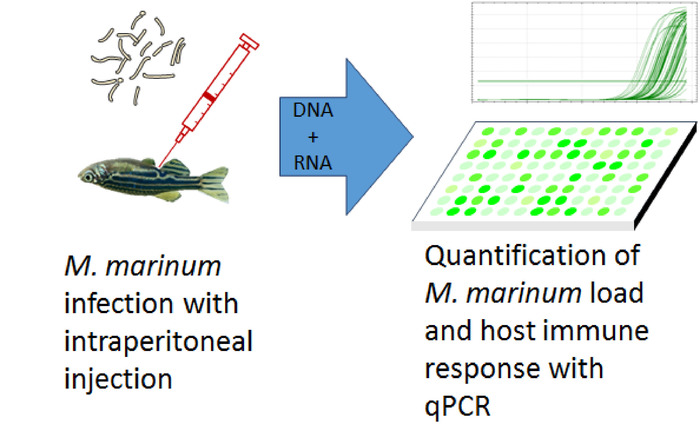
Figure 1: Workflow of studying the development of mycobacterial loads in the adult zebrafish. Adult zebrafish are infected with an intraperitoneal injection of M. marinum. DNA and RNA are extracted from the internal organs of the fish and the M. marinum load and host's immune responses are analyzed with quantitative polymerase chain reaction (qPCR). Please click here to view a larger version of this figure.
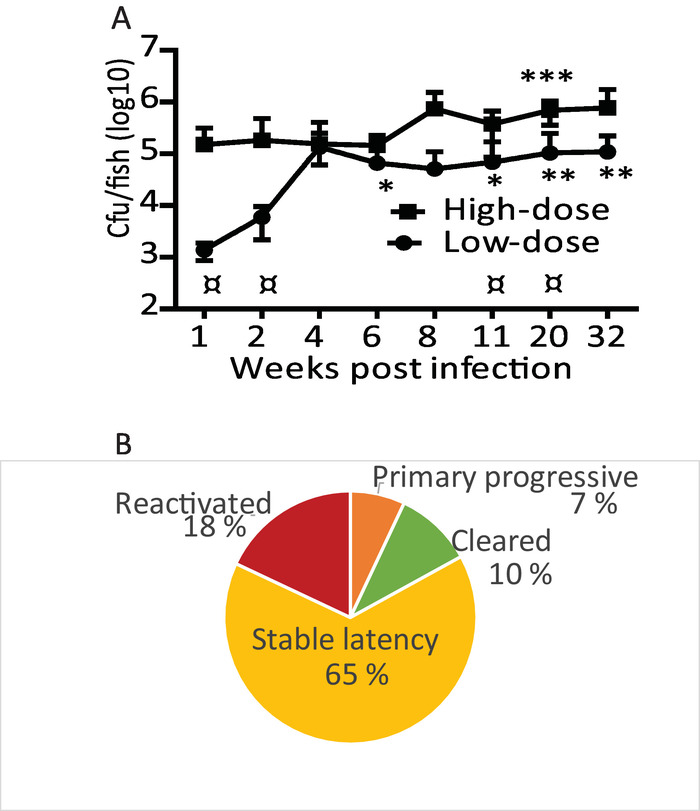
Figure 2: Injection of M. marinum into adult zebrafish causes a spectrum of disease states. (A) Zebrafish were injected with a low (34 ±15 CFU) or a high dose (2029 ±709 CFU) of M. marinum. Average loads for 5 fish (except 32 weeks high dose, n = 2) are shown with SD. Low-dose statistics: * p <0.05 compared with 1 week, ** p <0.05 compared with 1 and 2 week. High-dose statistics: *** p <0.05 compared with 1, 2, 8, 11 and 20 wk. ¤ low dose vs. high dose p<0.05. Modified from Parikka et al. 201219. (B) Typical distribution of disease outcomes within a wild-type zebrafish population infected with a low-dose of M. marinum. Please click here to view a larger version of this figure.
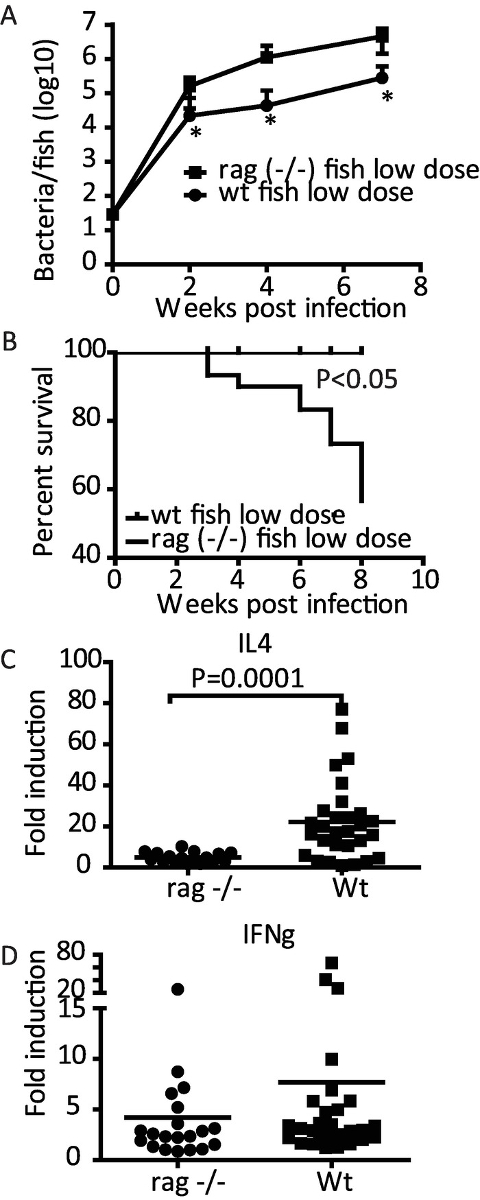
Figure 3: Adaptive immunity affects the course of mycobacterial infection in the adult zebrafish. Adult wild-type (wt) and rag1 (−/−) zebrafish were infected with a low dose (n = 30) of M. marinum. (A) The average mycobacterial loads were measured by qPCR at 2, 4, and 7 weeks post infection (wpi) (n = 10) *P<0.05. (B) The fish were euthanized upon the development of symptoms of the disease and survival plots were created. (C). The expression levels of il4 were measured at 4 wpi. (D) The expression levels of IFNγ were measured at 4 wpi. (A and B)modified from Parikka et al. 201219. (C and D) Modified from Hammarén et al. 201438. Please click here to view a larger version of this figure.
| Gene | Primer sequence | Annealing temperature |
| MMITS1 | F: CACCACGAGAAACACTCCAA | 65 |
| 16S–23SITS Locus AB548718 for M. marinum quantification | R: ACATCCCGAAACCAACAGAG | |
| loopern4 | F: TGAGCTGAAACTTTACAGACACAT | 61 |
| Expressed repetitive elements | R: AGACTTTGGTGTCTCCAGAATG | |
| il4 | GCAGGAATGGCTTTGAAGGG | 59.5 |
| ZDB-GENE-100204-1 | GCAGTTTCCAGTCCCGGTAT | |
| ifnγ1-2 | F: GGGCGATCAAGGAAAACGACCC, | 61 |
| ZDB-GENE-040629-1 | R: TAGCCTGCCGTCTCTTGCGT |
Table 1: Primer sequences and annealing temperatures. The sequences of the primers used and their optimized annealing temperatures. The primers for the M. marinum 16S-23S rRNA transcript have been optimized for a No-ROX qPCR kit and the other primers for a ROX including qPCR kit.
| Step | Time | Temperature |
| 1 | 3 min | 95 °C |
| 2 | 5 s | 95 °C |
| 3 | 10 s | 65 °C |
| 4 | 5 s | 72 °C (fluorescence detection) |
| 5 | Go to step 2. 39 times | |
| 6 | Melting curve analysis 55-95°C with 0.5°C intervals | |
| 7 | Forever | 4 °C |
Table 2: qPCR program for measuring M. marinum DNA. A qPCR protocol designed according to the manufacturer's instructions and optimized for measuring M. marinum DNA from zebrafish samples.
| Time | Temperature |
| 5 min | 25 °C |
| 30 min | 42 °C |
| 5 min | 85 °C |
| forever | 4 °C |
Table 3: cDNA synthesis program. Protocol for synthesizing cDNA from the extracted RNA of an infected zebrafish according to the manufacturer's instructions.
| Step | Time | Temperature |
| 1 | 30 s | 95 °C |
| 2 | 12 s | 95 °C |
| 3 | 30 s | Annealing °C |
| 4 | Go to step 2. for 39 times | |
| 5 | Melting curve analysis 65–95 °C with 0.5 °C intervals | |
| 6 | Forever | 4 °C |
Table 4: qPCR program for measuring host's gene expression. A qPCR protocol designed according to the manufacturer's instructions and optimized for measuring the expression of different zebrafish genes.
