Aggregated recombinant tau was covalently labelled with a pH-sensitive green dye. This dye dramatically increases its fluorescence upon its internalization in acidic organelles, thereby allowing for intracellular quantification. Labeled tau aggregates were incubated with anti-tau monoclonal antibodies. In particular, we used a chimeric version (mouse IgG1 Fc region) of CBTAU-28.1. This human antibody binds to the N-terminal insert region of tau and is able to bind in vitro generated tau fibrils13. In this assay, we also tested an affinity-improved version of CBTAU-28.1 – dmCBTAU-28.1. Fab fragments of CBTAU-28.1, in the parental and high-affinity mutant format, and a mouse IgG1 isotype control were used as controls.
BV-2 cells were incubated with the pre-formed immunocomplexes or aggregated tau alone for two hours in the presence of heparin to block antibody-independent tau uptake. After incubation, cells were trypsinized to remove the tau bound to the extracellular membrane and were analyzed for tau uptake by flow cytometry. As we recently described13, we observed that CBTAU-28.1 variants promoted uptake of tau in BV-2 cells in a dose-dependent manner. The uptake was Fc mediated since CBTAU-28.1 Fab fragments did not increase basal tau uptake (Figure 2). Moreover, the high affinity dmCBTAU-28.1 antibody mediated tau uptake into BV-2 cells to a higher extent than the wild-type antibody (Figure 2).
Antibody-mediated tau uptake and localization of tau aggregates in the endolysosomal compartment was confirmed by confocal microscopy (Figure 3) where the acidic cellular compartment was stained using a probe selective for low pH organelles. Intracellular puncta of green pH dye labeled tau aggregates were observed inside the cells that were incubated with CBTAU-28.1. Moreover, intracellular tau aggregates often colocalized with the low pH compartment selective red dye thus suggesting presence of tau aggregates in the acidic organelles. CBTAU-28.1 Fab fragments did not increase tau uptake again indicating an Fc-receptor mediated internalization mechanism (Figure 3).
Figure 1: Gating strategy used in flow cytometry analysis to detect tau internalization by BV-2 cells. Sample data from BV-2 only control (A-C), isotype control (D-F) and dmCBTAU-28.1 (G-I) are shown. BV-2 cell population was gated on a FSC-A vs SSC-A density plot excluding debris and dead cells (A, D, G). BV-2 cells were then further gated on a FSC-A vs FSC-H density plot to exclude cell doublets and aggregates (B, E, H). Single cell gate was used to generate a pH dye (FITC in these representative results) single parameter histogram (C, F, I) and determine geometric mean fluorescence intensity. Alternatively, percentage of pH dye-tau positive cells was calculated excluding negative cells as determined by using BV-2 only control. Please click here to view a larger version of this figure.
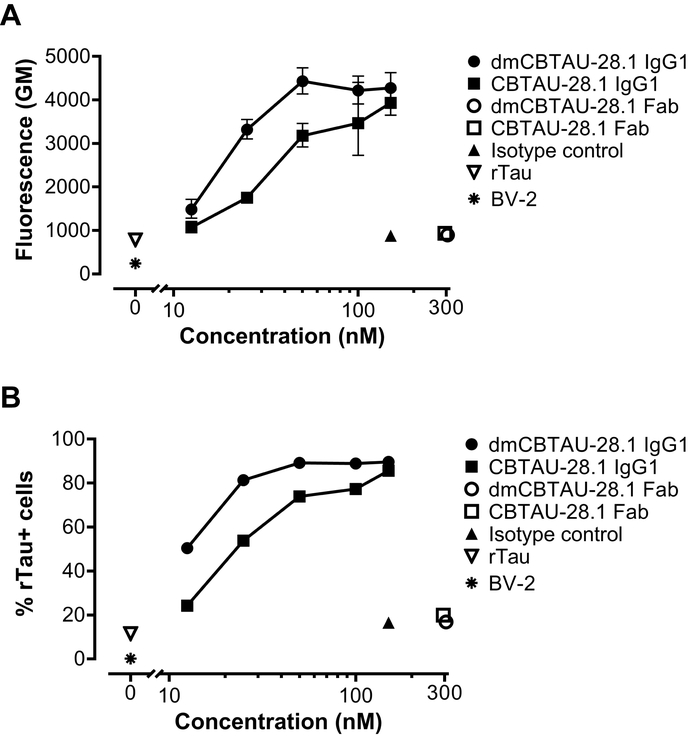
Figure 2: CBTAU-28.1 mediates uptake of tau aggregates into microglial BV-2 cells. Aggregated recombinant tau was covalently labelled with green fluorescence pH-sensitive dye and incubated with a mouse chimeric version of the human anti-tau antibody CBTAU-28.1, its affinity improved format, dmCBTAU-28.1, the corresponding Fab fragments, a mouse IgG1 isotype control antibody or no antibody (tau aggregates alone). Immunocomplexes were subsequently incubated with BV-2 cells for two hours in the presence of heparin to block antibody-independent tau uptake. Uptake of immunocomplexes was assessed by flow cytometry and expressed as the geometric mean (GM) of fluorescence intensity (A) or percentage of tau positive (tau+) cells (B). Error bars in (A) indicate the standard deviation of two independent experiments, while (B) shows a single experiment. Please click here to view a larger version of this figure.
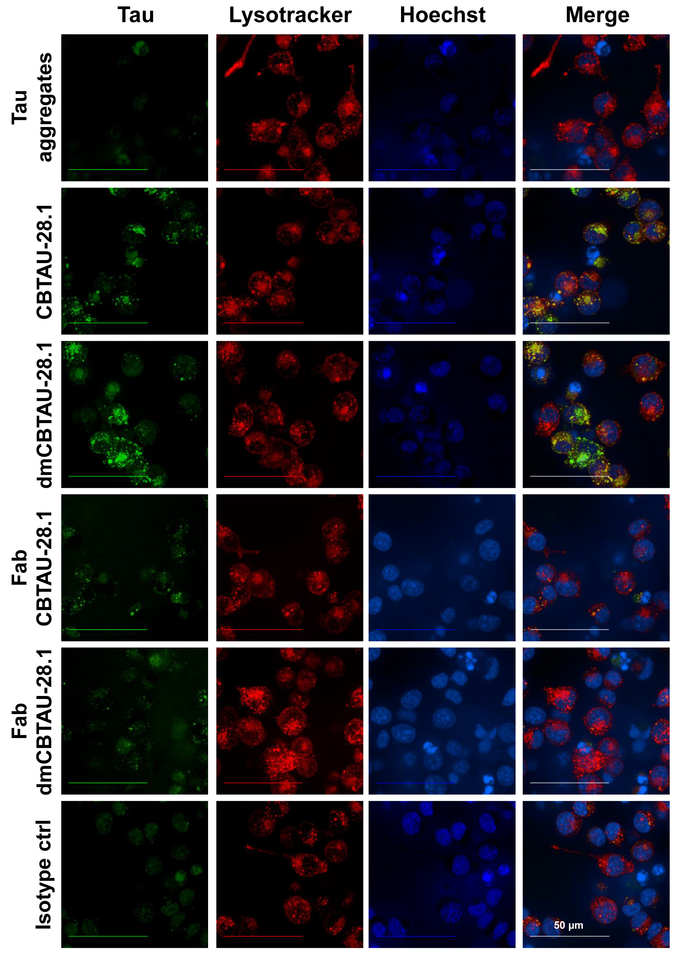
Figure 3: Tau aggregates are internalized by BV-2 cells and localize in cellular acidic organelles. Preformed tau-antibody immunocomplexes were incubated with BV-2 cells for two hours in the presence of heparin to block antibody-independent uptake. After incubation, nuclei were stained with a DNA specific blue dye and the acidic cellular compartment with a low pH compartment selective red dye. Live-cell imaging revealed intracellular puncta of labeled tau aggregates (green) inside the cells that were incubated with CBTAU-28.1 and dmCBTAU-28.1, but not with the isotype control. Moreover, intracellular tau aggregates often colocalized with the red dye (yellow) thus suggesting presence of tau aggregates in the acidic cellular compartment. CBTAU-28.1 Fab fragments did not increase tau uptake indicating an Fc-receptor mediated internalization mechanism. Images represent maximum intensity projections of a 20 planes Z-stack (0.5 µm planes) acquired with a 63X water immersion objective. Please click here to view a larger version of this figure.
