Figure 2 shows results from a digitonin titration experiment aimed to identify the proper amount of digitonin required for the extraction of SCs. This amount will vary depending on the tissue/cell type and whether the sample was frozen or not. For this experiment, a CIV in-gel activity was performed to visualize SCs isolated from fresh mouse liver mitochondria. Ratios from 2/1 to 10/1 g digitonin/g of protein were tested. The optimal amount of digitonin for this sample is 4 g/g, as it provides a good resolution of monomeric CIV, and high molecular weight SCs. At a lower ratio, bands are not clear and resolve into a smear during electrophoresis, whereas the use of higher ratio of digitonin leads to disruption of high molecular weight SC.
Figure 3 and Figure 4 show the results of a complete experiment performed on a preparation of mouse liver mitochondria extracted with 4 g digitonin /g protein. Proteins were separated using hybrid BN/CN-PAGE, standard BN-PAGE, or CN-PAGE. All three gels were casted at the same time and lanes were loaded with replicates of the same sample. Following electrophoresis, individual lanes were cut and processed for in gel activity measurement (CI, CII, CIV and CV on Figure 3) and immunoblotting (CI, CII, CIII, CIV, CV on Figure 4).
Addition of CB either momentarily in cathode buffer (i.e. hybrid CN/BN-PAGE) or in sample and cathode buffer throughout electrophoresis (i.e. BN-PAGE), considerably improves the mobility and resolution of SC bands, and individual respiratory complexes compared to CN-PAGE (Figure 3). Bands are easily distinguishable with the hybrid technique or BN-PAGE after in-gel activity for CIV, whereas in the same sample resolved by CN-PAGE, SCs and monomeric CIV reactive bands cannot be identified.
Figure 3 and Figure 4 show that the resolution and banding pattern of OXPHOS monomers and supramolecular assemblies is qualitatively comparable between hybrid CN/BN-PAGE and BN-PAGE. However, notable differences exist. First, the electrophoretic mobility of OXPHOS complexes is slightly reduced when proteins are separated using hybrid CN/BN-PAGE conditions vs standard BN-PAGE, due to reduced amount of CB. This mobility shift is greater for CIV monomers, followed by CV monomers, and CI (Figure 3 and Figure 4). Second, the blue background is lower in the hybrid CN/BN-PAGE compared to BN-PAGE (Figure 3, left lanes). As a result, high background levels following BN-PAGE completely masks the in-gel activity staining for CII, and enhances the background noise associated with the activity of CIV dimers (Figure 3). Third, the activity of CV is higher when samples are run under hybrid CN/BN-PAGE conditions compared to BN-PAGE (Figure 3), due to the reduced amount of CB, which is known to interfere with CV catalytic activity.26 CN/BN-PAGE also allows better preservation of CV supramolecular assemblies, as shown by a greater proportion of total CV activity being associated with CV dimers (Figure 3). Moreover, CV oligomers are visible under CN/BN-PAGE, while they are completely dissociated under BN-PAGE conditions. Interestingly, distinct bands displaying CV activity are also observed between CV monomers and dimers, when samples are run under CN/BN-PAGE (Figure 2).
Figure 5 shows a representative analysis of OXPHOS complex distribution in supramolecular assemblies. The image shows CI in gel activity of samples obtained from 4 distinct healthy mouse liver mitochondria preparations. Densitometry analysis allows to measure the area under the curve of CI-reactive bands, and to present the relative distribution of C1 activity in the monomeric (I1) and supramolecular forms (I1III2, I1III2IV1, I1III2IVn). Similar analysis can be performed following immunoblot.
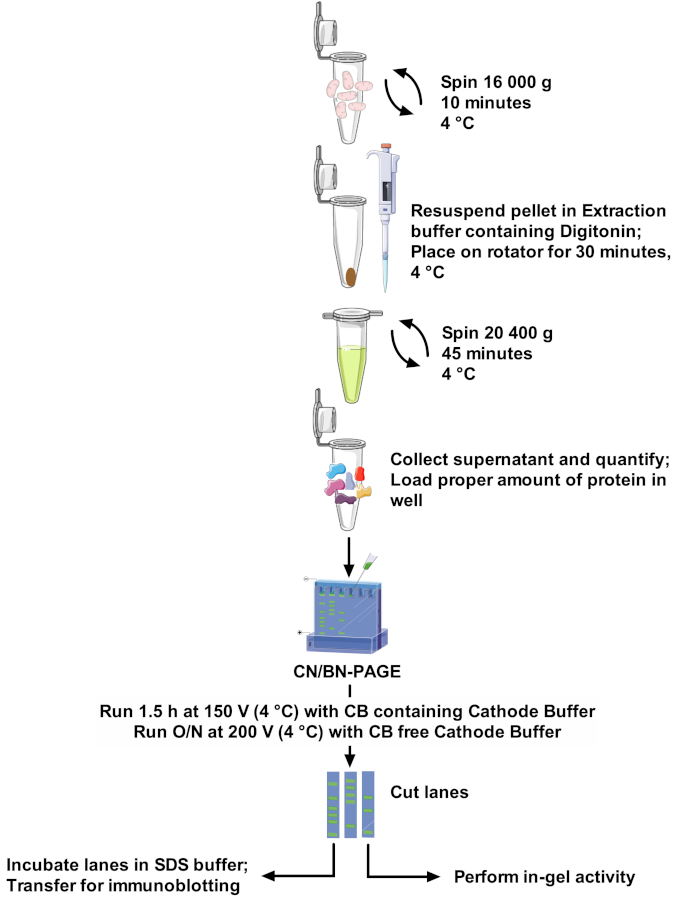
Figure 1: Assay workflow. Please click here to view a larger version of this figure.
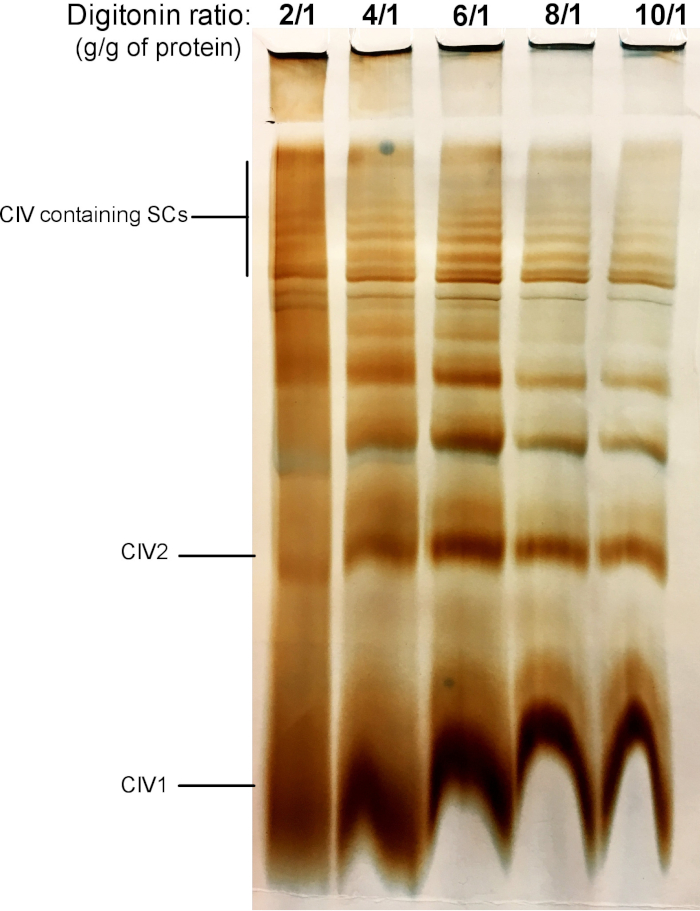
Figure 2: Digitonin titration to extract supercomplexes from fresh mouse liver mitochondria. This example shows aliquots of mouse liver mitochondria, isolated from one animal that was treated with increasing amounts of digitonin to extract respiratory supercomplexes. Samples were then resolved by hybrid CN/BN PAGE, and in-gel activity of CIV was determined. CIV1: complex IV monomers; CIV2: Complex IV dimers; SC: supercomplexes. Please click here to view a larger version of this figure.
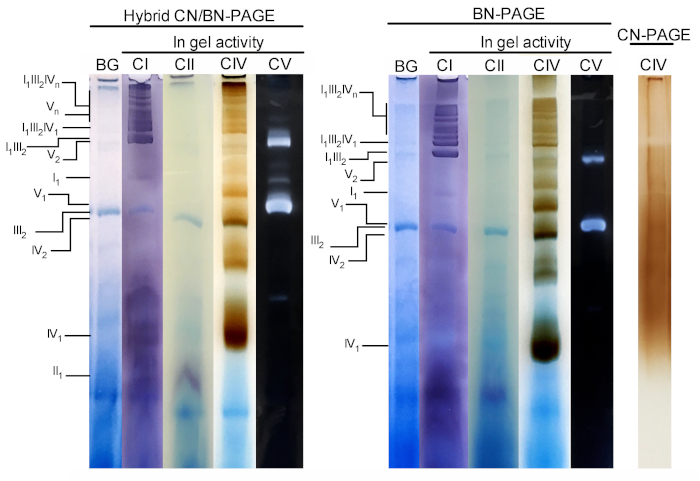
Figure 3: In-gel activity of OXPHOS complexes following hybrid CN/BN-PAGE, BN-PAGE or CN-PAGE. Liver mitochondria isolated from one mouse were treated with digitonin (4 g/g ratio digotonin/protein) to extract respiratory supercomplexes. Aliquots of this sample were then loaded on multiple wells in three distinct gels and submitted to CN/BN-PAGE, BN-PAGE or CN-PAGE. Each replicate lane within each gel was then cut and immediately used for in-gel activity assays (labeled CI, CII, CIV and CV). One lane was used as control to show background (labeled BG) staining with Coomassie Blue. OXPHOS complexes and supramolecular assemblies are identified using the standard nomenclature, with numbers in indices indicating the molecular stoichiometry of each OXPHOS complex. It should be noted that the position of CIII-containing supramolecular assemblies is based on immunodetection since in-gel activity for CIII was not performed in this particular experiment. Please click here to view a larger version of this figure.
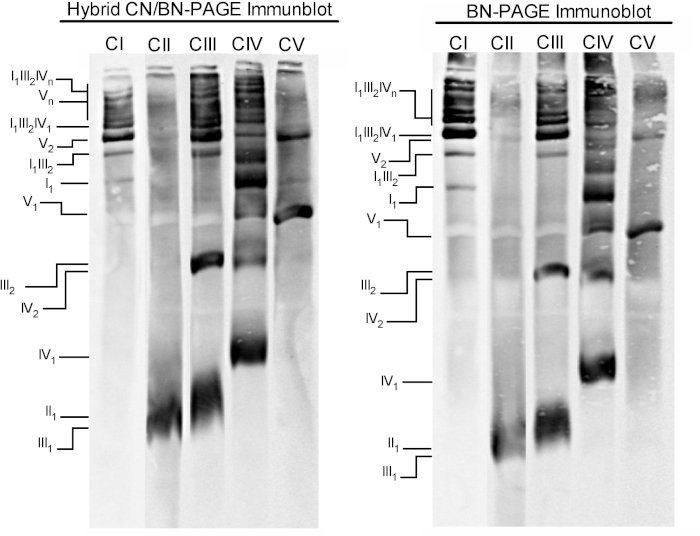
Figure 4: Immunoblot analysis of OXPHOS complexes following hybrid CN/BN-PAGE or BN-PAGE. Replicates from the experiments described in the Figure 3 legend were electro-transferred on a single membrane. After transfer, individual lanes were cut and incubated with specific antibodies recognizing CI, CII, CIII, CIV, and CV. OXPHOS complexes and supramolecular assemblies are identified using the standard nomenclature, with numbers in indices indicating the molecular stoichiometry of each OXPHOS complex. Please click here to view a larger version of this figure.
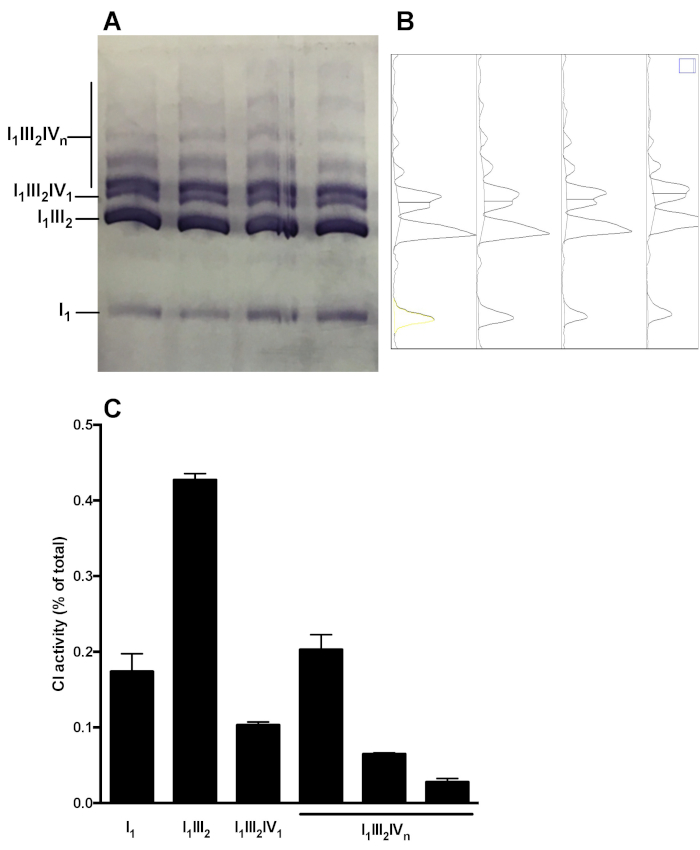
Figure 5: Quantification of CI distribution in monomeric and supramolecular assemblies. (A) CI in-gel activity determined following Hybrid CN/BN-PAGE of in liver mitochondria SC extracts obtained from 4 mice. (B) Densitograms obtained using ImageJ’s Gel Analysis Tool showing distinct peaks corresponding to CI monomers (I1) and various CI-containing supramolecular complexes (I1III2, I1III2IV1, and I1III2IVn). (C) quantification of the relative distribution of C1 activity. The data represent mean and SEM of the 4 mice. Please click here to view a larger version of this figure.
| Digitonin/protein ratio (g/g) | 2 g/g | 4 g/g | 6 g/g | 8 g/g |
| Volume of extraction buffer (µL) | 400 | 300 | 200 | 100 |
| Volume of 10 % stock digitonin (µL) | 100 | 200 | 300 | 400 |
| Total extraction buffer volume (µL) | 500 | 500 | 500 | 500 |
Table 1: Volumes required to extract SCs from 5 mg of mitochondrial proteins using various digitonin/protein ratios.
| Compound | Final Concentration |
| EDTA, pH 7.5 | 1 mM |
| HEPES | 30 mM |
| Potassium Acetate | 150 mM |
| Glycerol | 12% |
| 6-aminocaproic Acid | 2 mM |
Table 2: SC extraction buffer (final concentrations). Keep at 4 °C for a maximum of 3 months.
| Compound | Final Concentration |
| 3X Gel Buffer: Aliquot and keep at -20 °C, pH 7.5 | |
| Imidazole/HCl pH-7.0 | 75 mM |
| 6-aminocaproic Acid | 1.5 M |
| Acrylamide Buffer: Aliquot and keep at -20 °C | |
| Acrylamide | 99.5% |
| Bis-Acrylamide | 3% |
Table 3: Gel stock buffers.
| For 2 gels: | 4% (60 mL) | 12% (60 mL) | Stacking (4%) (25 mL) |
| 3X Gel Buffer | 19.8 mL | 19.8 mL | 8.25 mL |
| Acrylamide Buffer | 4.8 mL | 14.4 mL | 2 mL |
| H2O | 35 mL | 13.1 mL | 14.6 mL |
| Glycerol | – | 12 mL | – |
| APS 10% | 360 μL | 60 μL | 150 μL |
| TEMED | 24 μL | 12 μL | 10 μL |
Table 4: 4%–12% gradient gel.
| Compound | Final Concentration |
| Anode Buffer: Keep at 4 °C, pH 7.5 | |
| Imidazole | 25 mM |
| Cathode Buffer: Keep at 4 °C, pH 7.5 | |
| Tricine | 50 mM |
| Imidazole | 7.5 mM |
| With or without Coomassie Blue (G250) | 0.022% |
Table 5: Electrophoresis buffers.
| Compound | Final Concentration |
| Complex I activity Buffer: prepare fresh in 5 mM TRIS-HCl pH 7.4 | |
| Nitrotetrazolium blue | 3 mM |
| NADH | 14 mM |
| Complex II activity Buffer: prepare fresh in 5 mM TRIS-HCl pH 7.4 | |
| Succinate | 20 mM |
| PMSF | 0.2 mM |
| Nitrotetrazolium blue | 3 mM |
| Complex IV activity Buffer: prepare fresh in 50 mM Na-Phosphate pH 7.2 | |
| Cytochrome C | 0.05 mM |
| Diaminobenzidine | 2.3 mM |
| ATPsynthase activity Buffer: prepare fresh in water, adjust pH to 8 with KOH | |
| Glycine | 50 mM |
| MgCl2 | 5 mM |
| HEPES | 50 mM |
| CaCl2 | 30 mM |
| ATP | 5 mM |
Table 6: In-gel activity assay buffers.
| Compound | Final Concentration |
| Transfer Buffer | |
| Tris Base | 25 mM |
| Glycine | 192 mM |
| SDS | 4% |
| Methanol | 20% |
| TBST | |
| Tris Base | 20 mM |
| NaCl | 137 mM |
| Tween 20 | 0.1% |
Table 7: Immunoblotting buffers.
| Complex | Subunit | Clone |
| I | NDUFA9 | 20C11B11B11 |
| II | SDHA | 2E3GC12FB2AE2 |
| III | UQCRC2 | 13G12AF12BB11 |
| IV | COX4 | 1D6E1A8 |
| V | ATPB | 3D5 |
Table 8: Antibodies used for immunoblotting to detect respiratory chain SC. See Table of Materials for companies and lot numbers.
