A schematic of forming PSC colonies is depicted in Figure 1A. PSC spheroids are formed on a micro-fabricated plastic vessel for one day. As a representative result, 20,000 409B2 cells could be handled per 96-well of micro-fabricated plastic vessel, although other cell lines may require a higher density (30,000-40,000 cells per 96-well of micro-fabricated plastic vessel). Those spheres are spontaneously flattened to a nearly two-dimensional culture when plated onto a culture dish in the presence of LM511-E8.
A schematic of hemogenic induction is illustrated in Figure 1B. When PSC colonies grow to a diameter of 750 µm, the medium is sequentially changed to induce mesodermal organoids. The PSC colonies will gradually become a sunny side up structure during differentiation to mesodermal organoids by sequential medium change until Day 4. On Day 4, hemogenic endothelia are specified from mesodermal organoids by magnetic sorting and subsequently plated onto fibronectin in EHT medium. 5-10 million CD34+CD45– cells are expected from spheroids containing 1 million PSCs (Figure 1C).
It is observed that the cells morphologically change from endothelial to hematopoietic cells (Supplementary Video 1). A representative phase-contrast image of hematopoietic progenitor cells upon stimulation with hematopoietic cocktail on day 7 is shown in Figure 2A. Hematopoietic cell colonies emerged in the culture.
A representative flow cytometry plot is shown in Figure 2B. The whole culture is stimulated with hematopoietic cocktail and on day 7 is analyzed for the expression of CD34 and CD45 by flow cytometry. Hematopoietic progenitor cells express CD34 and CD45.
A CFU assay showed the generation of granulocytes/macrophage colonies from the CD34+ CD45+ hematopoietic progenitor cells (Figure 2C). The estimated colony number is 38 CFU-G/M per 10,000 cells (Figure 2D)
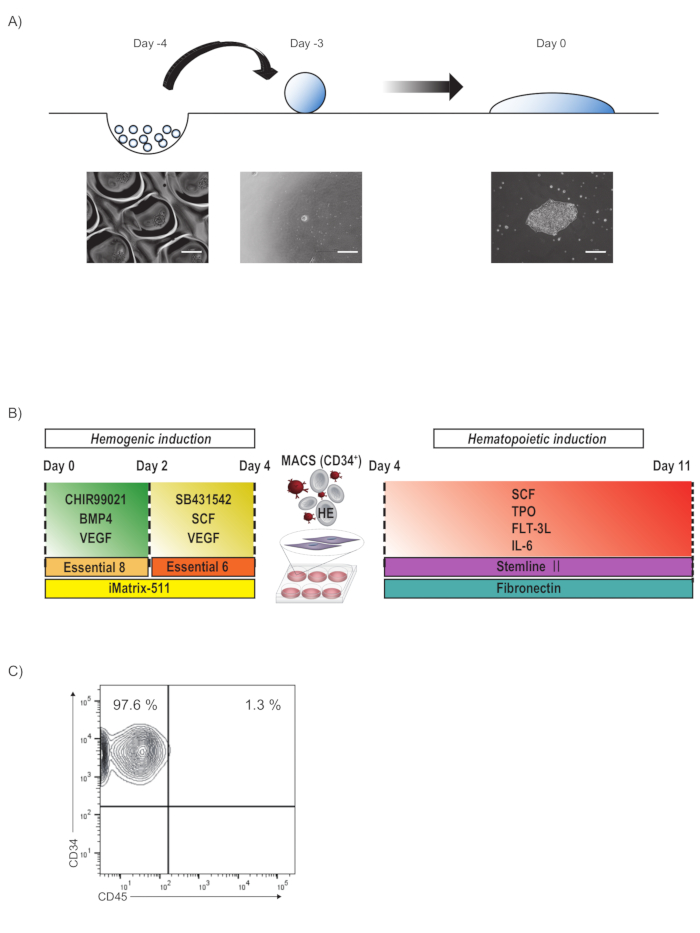
Figure 1: Schematic of tiling PSC colonies and subsequent hematopoietic differentiation via hemogenic endothelia. (A) Schematic process of forming PSC colonies. PSCs are maintained on a LM511-E8-coated 6-well plate in PSC maintenance medium. On Day -4, cells are detached and dissociated to the single-cell level using dissociating solution, subsequently plated onto micro-fabricated plastic vessel in PSC maintenance medium with Y-27632 and cultured overnight to form spheroids. On Day -3, spheroids are plated in PSC maintenance medium with LM511-E8 and cultured for three days. By Day 0, spheroids are spontaneously flattened to a nearly two-dimensional culture. Scale bars = 200 µm. (B) On Day 0, the medium is replaced with Day0 differentiation medium and cultured in the hypoxic incubator. On Day 2 of the differentiation, the medium is replaced with Day2 differentiation medium. On Day 4 of the hemogenic endothelium enrichment, CD34+ cells are magnetically sorted and cultured on a fibronectin-coated 12-well plate in EHT medium for 6 days. (C) Representative flowcytometry plot of CD45 and CD34 expression of day 4 cells sorted by CD34. Please click here to view a larger version of this figure.
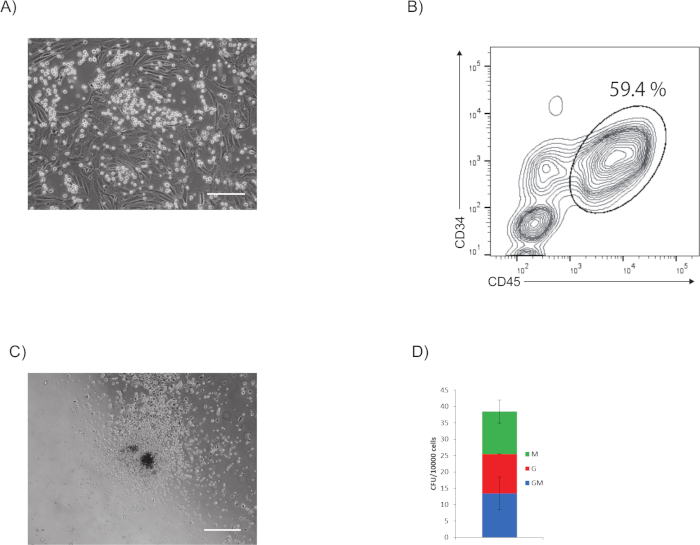
Figure 2: Analysis of hematopoietic property. (A) Representative phase-contrast image of hematopoietic progenitor cells after stimulation with hematopoietic cocktail on day 7. Scale bar = 200 µm. (B) Representative flow cytometry plot of CD45 and CD34 expression of whole culture upon stimulation with hematopoietic cocktail on day 7. (C) Representative phase-contrast image of a granulocyte/macrophage colony generated from Day 11 hematopoietic progenitor cells. Scale bar = 500 µm. (D) Number of CFUs per 10,000 cells. Please click here to view a larger version of this figure.
Supplementary video 1: EHT video. Please click here to download this file.
