A relatively fast and reliable method for the detection of total CoA in cultured cells and tissues has been developed by derivatizing the thiol of CoA to a fluorescent agent using mBBr, and then purifying the derivatized CoA-bimane using reverse phase HPLC. A standard curve is first generated, where known and increasing amounts of the CoA-bimane standard are injected individually and the areas under the peaks in the CoA-bimane chromatograms are plotted as a function of the input CoA-bimane (Figure 4). CoA-bimane has an absorbance maximum at λ393 nM and a representative HPLC profile shows the retention time of the CoA-bimane standard on a C18 HPLC column (Figure 3) using the elution program in Table 1. Representative standard curves of CoA-bimane, detected by measuring absorbance or fluorescence units, are shown in Figure 4A and Figure 4B, respectively. The standard curve in Figure 4A reflects the magnitude of absorbance of CoA-bimane at λ393 nm and Figure 4B represents the fluorescence of CoA-bimane (λex = 393 nm and λem = 470 nm) plotted versus the input amount of CoA-bimane. The CoA-bimane standard can be detected from 0.01 to 12,000 pmol and covers a 106-fold range when detection using both absorbance and fluorescence is combined. While the lower limit of detection of the standard is 0.01 pmol, the lower limit of CoA-bimane quantitation in experimental samples is 0.2 pmol which is about 5-fold greater than the baseline or background fluorescence in the chromatogram. The choice of detection by absorbance or fluorescence of CoA-bimane depends on the amount of CoA normally present in tissues or cells, together with the practicality of working with larger or smaller sample sizes. Absorbance is generally useful for tissue samples because handling 40-50 mg of starting material yields better recovery than 5 mg tissue samples, whereas fluorescence is generally useful for cultured cell samples which are smaller in size and there is greater confidence in interpolation of values at the lower end of the fluorescence standard curve. Laboratories with only absorbance detection may consider increasing or decreasing the starting sample size, increasing the HPLC injection volume or decreasing the volume for sample resuspension (Section 5.9 above) for measurements in cultured cells.
The conditions for hydrolyzing acyl-CoAs from tissues and cultured cells were optimized by adjusting the KOH concentration, and the time and temperature of subsequent incubation (data not shown). The optimum condition was found to be 0.25 M at 55 °C for 2 hours. The thiol group of free CoA plus any CoA liberated from thioesters were derivatized by reaction with mBBr following adjustment of the pH. Subsequent HPLC separation and typical detection profiles for mouse liver or human cultured C3A cells are indicated in Figure 5 as red peaks, with a retention time between 11 and 12 minutes using the elution program described in Table 1. Typical amounts of biological starting material are 30-40 mg of murine liver (wet weight), 6-8 x 106 cells for human "liver-like" C3A cells, or ~1.3 x 107 cells for human HEK293T cells. The C3A cells were treated with a PanK experimental drug PZ-2891 at 10 uM which elevates CoA17 (Figure 6). The CoA measurements in HEK293T cells when PanK isoforms are overexpressed show CoA measurements over a wide range (431-6925 pmoles/µL) (Figure 6). The sample sizes required for this methodology are practical for application to many experimental contexts.
The area under the CoA-bimane peak was calculated using software provided with the HPLC. The peak limits can be determined automatically by the HPLC software pending proper adjustment of input values for baseline correction and peak definition, but our laboratory prefers to manually designate the CoA-bimane peak in each chromatogram, particularly for unfamiliar biological samples.
| Time (min) | Flow Rate (mL/min) | % A | % B | Curve |
| 0 | 0.5 | 90 | 10 | 0 |
| 2 | 0.5 | 90 | 10 | 6 |
| 6 | 0.5 | 85 | 15 | 6 |
| 18 | 0.5 | 60 | 40 | 6 |
| 23 | 0.5 | 60 | 40 | 6 |
| 25 | 0.5 | 90 | 10 | 6 |
| 30 | 0.5 | 90 | 10 | 66 |
Table 1: HPLC Program for CoA-bimane Separation. Buffer A: 50 mM KH2PO4, pH 4.6; Buffer B: Acetonitrile. The mixing of Buffer A and Buffer B follows Curve 6 which is a linear gradient, with an intervening Curve 8 which is a medium concave gradient.
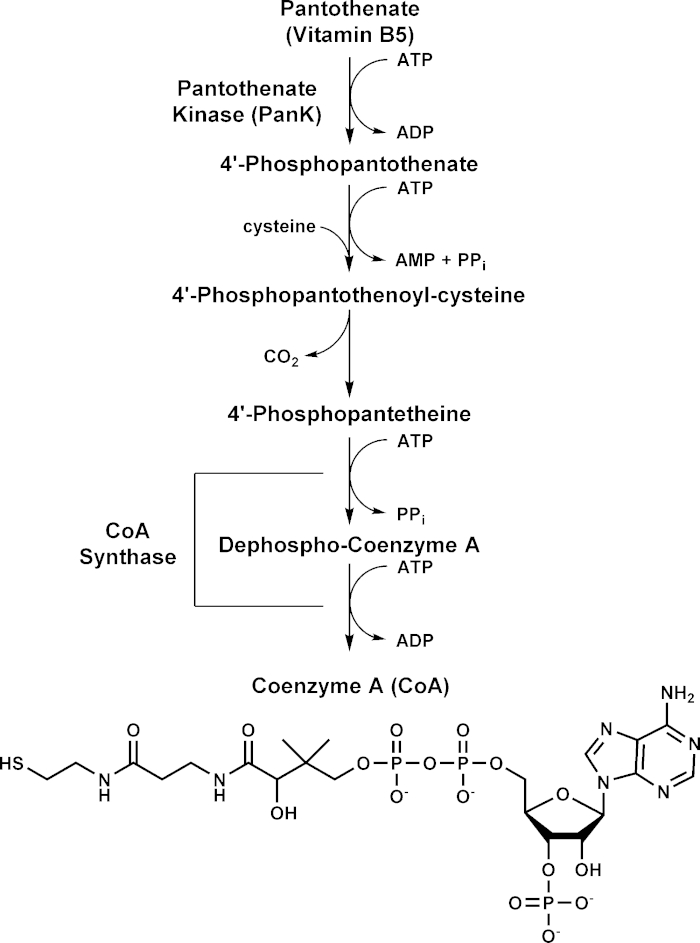
Figure 1. Coenzyme A biosynthesis pathway. Pantothenate kinase (PanK) catalyzes the phosphorylation of pantothenate (vitamin B5) to 4′-phosphopantothenate in the first step of CoA biosynthesis. Formation of phosphopantothenate is followed by condensation with cysteine catalyzed by 4′-phosphopantothenoylcysteine synthase and then decarboxylation to form 4′-phosphopantetheine by 4′-phosphopanthenoylcysteine decarboxylase. 4′-Phosphopantetheine is converted to Coenzyme A (CoA) in a two-step process catalyzed by CoA Synthase. Please click here to view a larger version of this figure.
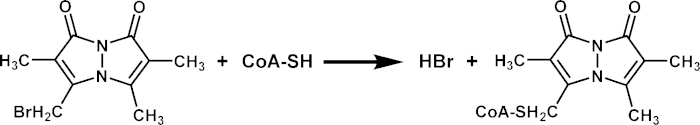
Figure 2. mBBr Reaction with CoA. Monobromobimane (mBBr) is mixed with CoA-SH (free CoA) and incubated at room temperature for 2 hours in the dark. Non-fluorescent mBBr becomes fluorescent when bound to CoA to form CoA-bimane. Please click here to view a larger version of this figure.
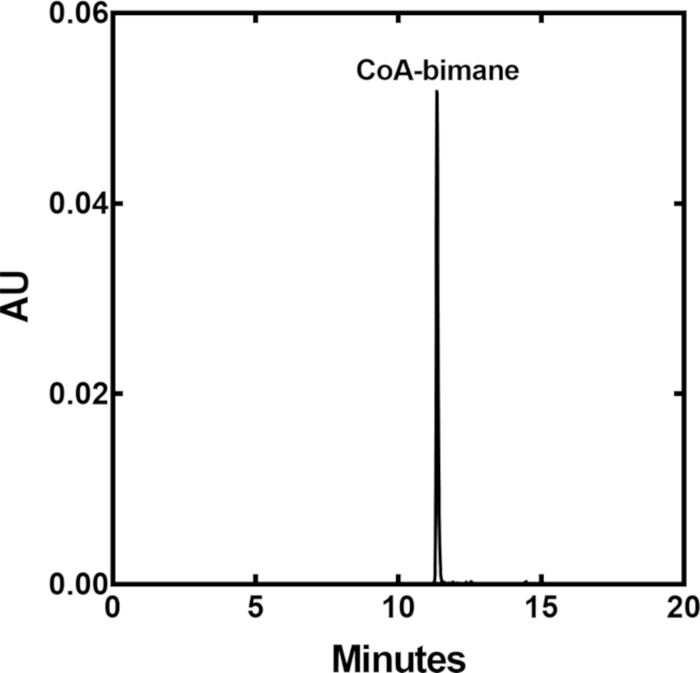
Figure 3: Absorbance HPLC Trace of CoA-bimane Standard. CoA-bimane was separated on a Gemini C18 column using the HPLC program in Table 1. Typical retention time (min) is indicated by aborbance units (AU). Please click here to view a larger version of this figure.
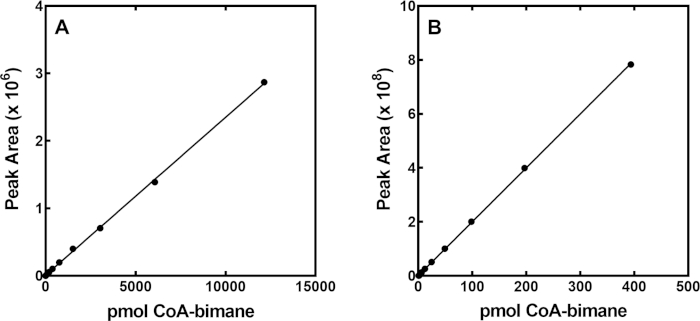
Figure 4: CoA-bimane standard curves detected by absorbance or fluorescence. (A) The standard curve measured using absorbance detection units for CoA-bimane was measured at λ393 nm. Tissue samples are usually evaluated using the absorbance standard curve. (B) The standard curve measured using fluorescence detection units for CoA-bimane at λex = 393 nm, λem = 470 nm. Cultured cells are usually evaluated for CoA content using the fluorescence standard curve. Duplicate standards (and samples) are routinely evaluated. Please click here to view a larger version of this figure.
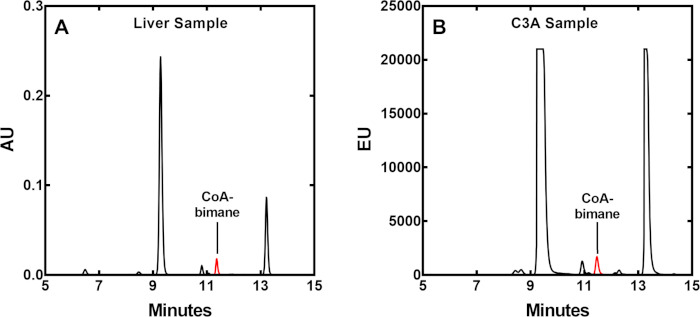
Figure 5: HPLC traces of typical liver or cultured cell samples. (A) CoA-bimane identification and quantification of content in mouse liver extract. Absorbance units (AU) are indicated. (B) CoA-bimane identification and quantification of content in HepG2/C3A cultured cells. Fluorescence emission units (EU) are indicated. The CoA-bimane peaks are shown in red. Please click here to view a larger version of this figure.
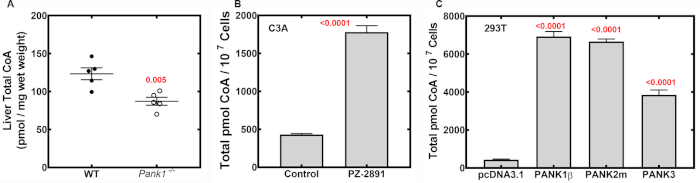
Figure 6. CoA levels in mouse liver, cultured C3A and HEK293T cells. (A) CoA measurement in mouse liver from pantothenate kinase knockout (Pank1-/-) and matched wild-type (WT) animals. Pank1-/- animals have lower CoA in the liver compared to WT animals due to the absence of Pank1 expression9. The data were obtained using 5 male mice per genotype and are represented as the mean ± SEM. (B) CoA levels in HepG2/C3A cells treated with either dimethylsulfoxide (vehicle control) or PZ-2891, an experimental drug that modulates PanK activity at 10 uM17. The cellular CoA level is elevated following 24-hour incubation with PZ-2891. (C) CoA levels in HEK293T cells transfected with either empty vector pcDNA3.1, or cDNAs encoding human PANK isoforms: PANK1β, PANK2m which is the mature, processed isoform of human PANK2, or PANK3. CoA levels are elevated by overexpression of all active PANK isoforms. The data in (B) and (C) are from independent triplicate samples and are represented as the mean ± SEM. The statistical analysis was done using the unpaired t-test and the values of significance are shown in red. The data in panels (B) and (C) are adapted from Sharma et al.12 Please click here to view a larger version of this figure.
