Figure 5 shows the development of disease symptoms of tomato plants with roots transformed with an empty vector (EV), and plants with roots transformed with an RNAi construct targeting SlCESA6 (Solyc02g072240). The disease index data (Figure 5A) are collected from the same experimental unit (each plant) over time according to an arbitrary scale from 0 to 4, and do not follow a Gaussian distribution, ruling out the use of standard tests for parametric data. Moreover, there is an intrinsic plant-to-plant variation in this kind of experiments, due to the colonization and infection dynamics. Below we considered different methods for the representation and statistical analysis of symptoms associated to Ralstonia infection.
As a standard approach, a U Mann-Whitney two-tailed non-parametric test to compare both control (EV) and SlCESA6-RNAi infection curves was used. According with this analysis, the difference between the medians of both curves appears to be non-significant (P = 0.16).
It is also possible to quantify the area under the disease progress curve (AUDPC), which allows combining multiple observations of disease progress into a single value. AUDPC showed a higher value for control plants (EV; 28.75 ± 4.75) compared to SlCESA6-RNAi (21.01 ± 4.74) plants at the end of the infection process, indicating that SlCES6-silenced plants are more resistant to Ralstonia infection than control plants (Figure 5B).
Confidence intervals (CI) offer a way of estimating, with high probability, a range of values in which the population value (or parameter) of a given variable is found. As it is shown in Figure 5C, the area of 95% CI for control (EV) and SlCESA6-RNAi infection curves estimate a higher chance of resistance when SlCESA6 is silenced.
Disease index values can be transformed into binary data, considering a disease index lower than 2 corresponding to "0", and a disease index equal or higher than 2 corresponding to "1"12. This allows the representation of a survival curve after Ralstonia inoculation. This transformation is based on the observation that, once the plants start developing clear symptoms (disease index of 2), they are considered as "infected" and will die as a consequence of this infection. The differences in the survival rate between EV and SlCESA6-RNAi plants were not statistically significant according to a Gehan-Breslow-Wilcoxon Statistical Test (P = 0.13) (Figure 5D).
In addition to performing statistical analysis, and regardless of the obtained P values, it is worth interpreting these data based on the reproducibility of the observed tendencies within different replicates (Supplementary Figure 1). In keeping with this notion, an increasing number of scientists have recently commented on the risks of overusing strict statistical thresholds (such as the standard P ≤ 0.05)16.
The expression of SlCESA6 was analyzed in two randomly selected transformed roots before the inoculation step, showing that the enhanced resistance to Ralstonia correlates with the reduced expression of SlCESA6 (Figure 5E). Using this method, a transformation rate of 35−40% after two rounds of selection can be obtained (Figure 5F). This value can be increased by performing additional selection rounds5.

Figure 1: Plant preparation, transformation and selection. (A) Germination of tomato seeds in half-strength MS (1/2) medium. (B) Germinated tomato seeds placed on filter paper lying on a plate containing 1/2 MS medium. Tomato seedlings before (C) and after (D) cutting the radicle and the bottom of the hypocotyl. (E) Tomato seedling transformation by dipping in bacterial biomass. (F) Transformed tomato seedlings covered with filter paper to maintain humidity. Emergence (G) and removal (H) of new (non-transformed) hairy roots. (I) Transformed roots. (J) Selection of transformed tomato hairy roots by visualization of DsRed fluorescence. Red arrows indicate positive transformed roots expressing DsRed fluorescence. (K) Development of the transformed roots as the main roots. (L) Visualization of DsRed fluorescence in mature transformed roots. Please click here to view a larger version of this figure.

Figure 2: Alternative method based on antibiotic selection. (A) Half-filled square plate containing 1/2 MS medium. (B) Cut seedlings disposed on filter paper lying on 1/2 MS medium with antibiotic, after removal of first non-transformed hairy roots. (C) Roots covered with additional filter paper. (D) Tomato transformed roots grown on 1/2 MS medium with antibiotic. (E) Seedlings transformed with pK7GWIWG2_II-RedRoot (kanamycin 50 µg/mL), expressing DsRed, as positive control. Please click here to view a larger version of this figure.

Figure 3: Transfer of transformed tomato to inoculation pots. (A) Root-transformed tomatoes transferred to full-soaked inoculation pots. (B) Transformed tomatoes in inoculation pots covered with a plastic lid to maintain high level of humidity. (C) Two-to-three-week-old root-transformed tomatoes, after transference to inoculation pots, ready to be inoculated. (D) Three-week-old tomato plants after removing the soil. (E) DsRed fluorescence in roots of 3-week-old transformed tomato plants. Please click here to view a larger version of this figure.

Figure 4: Ralstonia soil-drenching inoculation. (A) Materials needed for R. solanacearum GMI1000 inoculation. (B) Soil-drenching of transformed tomatoes in inoculation pots with R. solanacearum GMI1000 inoculum. (C) Inoculated tomatoes placed on a layer of potting soil. (D-H) Ralstonia disease symptoms scale ranging from 0 (no symptoms) to 4 (complete wilting). Please click here to view a larger version of this figure.
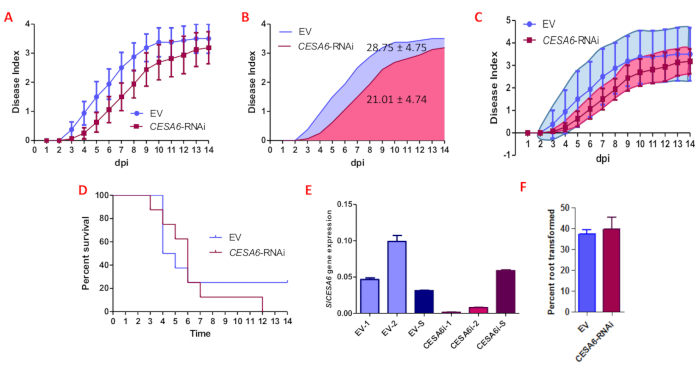
Figure 5: SlCESA6 silencing enhances resistance to R. solanacearum. (A) Disease symptoms of RNAi-mediated SlCESA6-silenced (CESA6-RNAi) and empty vector (EV) root-transformed tomato plants upon inoculation with R. solanacearum. Values correspond to the means ± SE of eight plants. (B) Area under the disease progress curve of plants shown in panel A. (C) 95% confidence Interval of plants shown in panel A. (D) Percent of surviving plants shown in panel A. (E) Gene expression analysis by qRT-PCR of RNAi-mediated SlCESA6-silenced (CESA6i) and EV root-transformed tomato plants in roots (1,2) and shoot (S). Values correspond to the means ± SE of three technical replicates. (F) Transformation rate of EV and CESA6-RNAi root-transformed tomato plants of two independent experiments. Values correspond to the means ± SE, after two rounds of selection. Please click here to view a larger version of this figure.
| Primer name | Primer sequence (5'-3') |
| EFα-1-F | GGTGGCGAGCATGATTTTGA |
| EFα-1-R | CGAGCCAACCATGGAAAACAA |
| qCESA6-F | GATCTGGTTCGCTTTCTCGT |
| qCESA6-R | TCCCTCCCTTTCATACCTTG |
| CESA6-RNAi-F | CACCGGCGAACAAGTGGGGTTAG |
| CESA6-RNAi-R | TTTGAGACTTTGGCACTGGA |
Table 1: Primer sequences.
| Ingredients | For 1 L |
| Bacto peptone | 10 g |
| Yeast extract | 1 g |
| Casamino acids | 1 g |
Table 2: Phi (Ø) medium composition.
Supplementary Figure 1: Reproducibility of the result shown in Figure 5A. Disease symptoms of RNAi-mediated SlCESA6-silenced (CESA6-RNAi) and EV root-transformed tomato plants upon inoculation with R. solanacearum. Values correspond to the means ± SE of eight plants. Gene expression analysis was performed by qRT-PCR of RNAi-mediated SlCESA6-silenced (CESA6i) and Empty Vector (EV) root-transformed tomato plants in roots (1,2) and shoot (S). Values correspond to the means ± SE of three technical replicates. Please click here to download this figure.
