The protocol described above allows for the preparation of viable slices from the septal pole of the dorsal hippocampus in mature rats. A key factor in this protocol is the perfusion of chilled sucrose-ACSF, prior to slice preparation, resulting in healthy CA1 PCs proximal to the slice surface. The quality of the slice produced is assessed visually under IR-DIC optics, and healthy cells identified as having large, ovoid-shaped cell bodies are located throughout the full extent of stratum pyramidale, from the compact layer, into stratum oriens (Figure 2A, black arrow). Unhealthy slices are identified as having dead cells on the surface and rarely in the depths of the slice (Figure 2A, red arrow), which are identified on the basis of having either condensed and highly contrasted somata, or large “ballooned” somata with condensed nuclei.
Confirmation of slice health is achieved by performing whole-cell patch clamp recordings from putative healthy neurons. Whole-cell patch-clamp recordings are achieved with rapid, spontaneous gigaohm seal formation (15.5 ± 2.9 s; Figure 2B), comparable to those previously reported31. When the membrane is ruptured, healthy neurons in mature rats possess hyperpolarized resting membrane potentials (Mean: -65.6 ± 1.5 mV, Range: -55.6 to -73.9 mV; 15 PCs from 4 rats) and relatively low input resistances (Mean: 90.3 ± 5.2 MΩ, Range: 54.9 to 134.2 MΩ; 19 PCs from 4 rats). General slice quality is confirmed by high-fidelity spontaneous EPSCs (Figure 2C,2D), given low electrical noise (<10 pA peak-to-peak) when filtered at 10 kHz. Furthermore, stable cell recordings of hyperpolarized neurons require typically a <200 pA holding current, which is stable over long periods, due to the absence of network activity in the submerged recording conditions of this slice preparation.
Whole-cell patch clamp recordings from dorsal CA1 PCs allow for direct measurement of action potential discharge properties. Provided that the slice quality is sufficiently high, many cells can be recorded from a single slice within a short time frame (~1 hour). A key determinant of cell viability is the presence of an intact dendritic tree, and the axon surviving beyond the initial segment. The slicing angle of 10° from vertical allows for the preservation of this cellular anatomy, with cells preserved within the plane of slicing (Figure 3A). Healthy CA1 PCs from adult rats typically have a hyperpolarized membrane potential of -60 mV to -70 mV, input resistances of 100-200 MΩ and membrane time-constants of 20-40 ms when measured at the soma (Figure 3B). A key requirement for neuron inclusion in datasets is the presence of action potentials in response to depolarizing stimuli. CA1 PCs in adult rats present increasing numbers of action potentials to depolarizing stimuli, from the rheobase current to the maximum tested currents (400 pA), at which trains of action potentials display both adaptation of inter-spike times and accommodation of action potential amplitude (Figure 3C). The use of a variable frequency sinusoidal wave (0.1 -20 Hz over 20 s) allows for characterization of the membrane resonance of the recorded neurons (Figure 3D). Finally, temporally controlled trains of action potential discharge over a range of frequencies allow for comparison of accommodation and recruitment of K+ channels associated with the resulting hyperpolarization (Figure 3E). Following post-hoc confirmation of intact dendrites using streptavidin visualization of biocytin labelling performed during recordings, the spontaneous EPSC frequency measured from the continuous recording (Figure 2B, upper) allows for characterization of CA1 PC integration into the local network.
In summary, optimization of slice quality of the dorsal extent of the hippocampus allows for whole-cell recordings from multiple neurons per slice. This slice preparation facilitates the collection of large datasets of intrinsic excitability, the establishment of intra-animal variability measures, and the production of slices of sufficient quality to perform paired recordings from synaptically-coupled neurons.
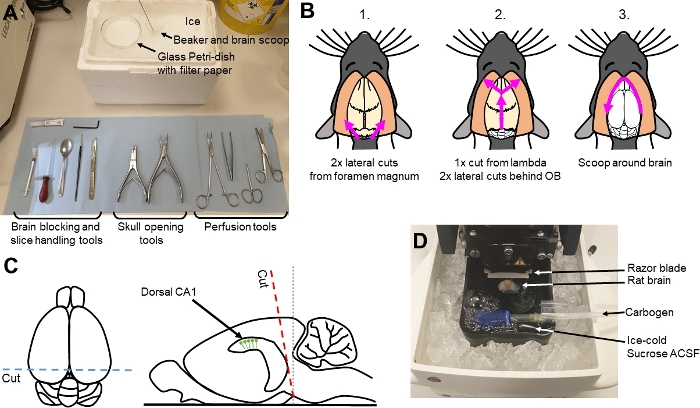
Figure 1: Overview of experimental setup and dissection schematic. (A) Experimental tools for all aspects of slice preparation, labelled according to use. (B) Cartoon depicting the directions of cuts with bone snippers and the movement of the spatula (pink arrows) to remove the brain from the skull. (C) Overview of the cutting angle (dashed red line) to allow preservation of dorsal CA1. (D) Overview of the slicing chamber with the brain mounted, anterior aspect facing up. Please click here to view a larger version of this figure.
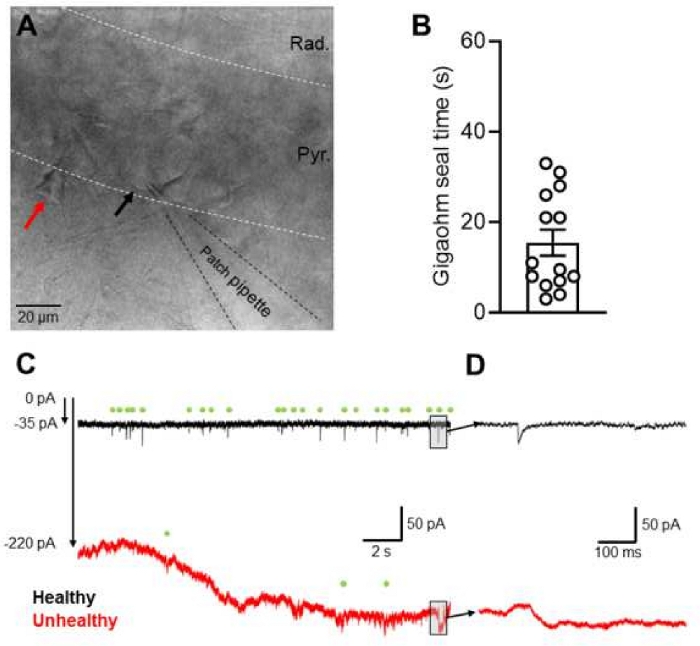
Figure 2: Identification of healthy neurons from the CA1 region of dorsal hippocampus. (A) Micrograph of area CA1 from an acute dorsal hippocampal slice, produced from a near coronal brain slice. The patch pipette is shown in a whole-cell configuration from a healthy neuron in the slice (indicated with black arrow). A nearby highly contrasted neuron to be avoided for recording is indicated (red arrow). (B) Representative continuous recordings of spontaneous EPSCs performed at -70 mV voltage-clamp from a stable recording of a healthy CA1 PC (black), with spontaneous EPSCs identified (green circles) and an unstable/unhealthy cell recorded under the same conditions (red). The holding current required to maintain -70 mV voltage-clamp is indicated. (C) Expanded view from the region of the trace in (B) indicated with a shaded box. Note the EPSC present in the top, stable trace (black) and the unstable, noisy trace (red). Please click here to view a larger version of this figure.
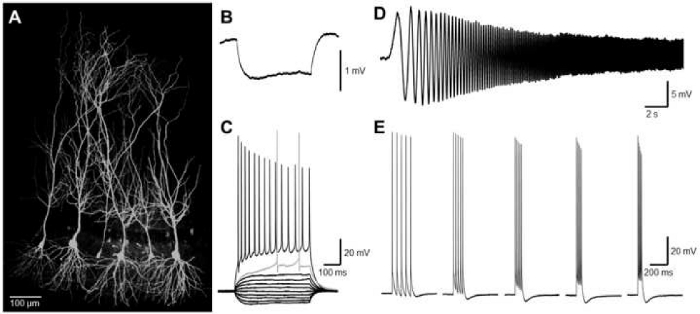
Figure 3: Cell identification and intrinsic electrophysiology, as measured by whole-cell patch-clamp recording from dorsal hippocampal CA1 PCs. (A) Visualization of biocytin with fluorescent-conjugated streptavidin labelling, followed by confocal imaging, from a slice containing multiple (6) CA1 PCs recorded sequentially, confirming the cellular identity of neurons recorded. (B) Average response to a -10 pA, 500 ms small hyperpolarizing step to ascertain passive membrane properties. (C) Voltage response of an identified CA1 PC to hyper- to depolarizing current steps (-100 to +400 pA, 500 ms duration). Action potential discharge is shown at both the rheobase current (grey sweep) and maximal discharge at 400 pA. (D) Membrane response to a 100 pA sinusoidal wave, frequency modulated from 0.1 – 20 Hz. Note the larger voltage response at the lowest cycle rates. (E) Trains of action potentials generated in response to trains of 5 stimuli (2 nA, 2 ms duration) over a range of frequencies (indicated). Please click here to view a larger version of this figure.
| Solution | Composition (in mM) | Notes | |
| Sucrose-ACSF | 87 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 glucose, 75 sucrose, 7 MgCl2, 0.5 CaCl2 | Chill before use | |
| Recording-ACSF | 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 glucose, 1 MgCl2, 2 CaCl2 | Pre-warm before use | |
| Intracellular Solution | 142 K-gluconate, 4 KCl, 0.5 EGTA, 10 HEPES, 2 MgCl2, 2 Na2-ATP, 0.3 Na2-GTP, 1 Na2-Phosphocreatine, 2.7 Biocytin (Osm ≈ 300 mOsm) | pH to 7.35 with 10 M KOH | |
| (K-gluconate) | |||
Table 1: List of solutions used in the preparation and recording of brain slices. Solutions are listed with their components reported as mM concentration. Specific notes prior to use are listed.
