The SDS-PAGE profile of the Ni affinity purification of MOMP-tNLP from a 1 mL cell-free reaction is shown in Figure 1B. The reaction resulted in high levels of expression for both the MOMP and the Δ49ApoA1 protein. Previous results showed that the cell-free expression of Δ49ApoA1 in the presence of DMPC and telodendrimer resulted in the formation of telodendrimer nanolipoprotein particles (tNLPs)4. The co-elution of MOMP with Δ49ApoA1 indicated that MOMP is associated with tNLPs, as the His-tag is only present on the tNLP scaffold Δ49ApoA1 and not on MOMP. MOMP is a highly insoluble protein that can only be eluted through complexing with tNLPs, which have been shown to facilitate the solubilization of membrane proteins.
The elution fractions containing MOMP-tNLPs were pooled and the total protein concentration determined using a fluorescence-based quantitation device, or a device that measures concentration through absorbance at 280 nm, following the manufacturer's instructions for protein quantitation. To allow for precise dosing of the MOMP vaccine, it is also important to determine the concentration of MOMP in the purified complexes. We developed a method to quantify MOMP based on gel densitometry (Figure 2) wherein a purified recombinant MOMP with known concentration was used as the standard. By establishing the standard curve and comparing it to the MOMP-tNLP sample, the MOMP concentration can be quantified accurately. The determination of MOMP concentration in the purified sample enabled the estimation of the yield of MOMP in cell-free reactions at various scales, which is important for planning the reaction setup appropriate to downstream studies (Table 3).
MOMP needs to form oligomers to elicit a robust immune response11. To test the oligomeric state of MOMP, MOMP-tNLP was analyzed in the presence and absence of both heat and the reducing agent dithiothreitol (DTT, 50 mM, Figure 3A). Higher-order oligomers of MOMP were identified through SDS-PAGE when samples were not treated with heat and DTT. In comparison, samples treated with heat in the presence of DTT showed primarily two distinct bands on the gel, corresponding to MOMP and Δ49ApoA1 (approximately 40 kDa and 22 kDa, respectively). These results closely resemble the gel banding pattern attributed to oligomer formation of MOMP, which is critical to its effectiveness.
Further western blot analysis using MAb40, an antibody against the linear epitope on the variable domain of MOMP protein, showed a similar banding pattern, confirming the oligomer formation by MOMP protein in its non-denatured state (Figure 3B). An important factor impacting MOMP oligomer formation is the ratio between the MOMP plasmid and the Δ49ApoA1 plasmid during the cell-free reaction setup. Table 4 lists the ratio of plasmids and the resulting insertion rate of MOMP into tNLPs. Previous studies indicated that chlamydial MOMP and other outer membrane proteins may exist primarily as trimers12. To maximize the trimer formation in the cell-free reaction, it is desirable to have the insertion rate close to three MOMP proteins per NLP, which corresponds to a ~25:1 MOMP-to-Δ49ApoA1 plasmid ratio.
A dot blot assay was used as a more streamlined method to detect the presence of MOMP and tNLP. The MAb40 antibody was used to detect total MOMP. The MAbHIS antibody targeted to the His-tag on the Δ49ApoA1 scaffold of the tNLP was used to assess the presence of tNLP. The co-signaling of MAb40 and MAbHIS antibodies indicated MOMP-tNLP formation. The control reaction produced empty tNLP, which only showed a positive signal from MAbHIS (Figure 3C). To test the immunogenicity of MOMP-tNLPs produced in the cell-free reaction, we adjuvanted MOMP-tNLP with CpG + FSL-1 and injected intramuscularly (i.m.) into mice in a prime-boost regimen as described above. Sera were collected from the immunized mice, and MOMP-specific IgG antibody was measured using a western blot assay (Figure 4). The sera from mice injected with adjuvanted MOMP-tNLP showed strong MOMP binding, indicating that MOMP-tNLP could elicit an immune response in vivo.
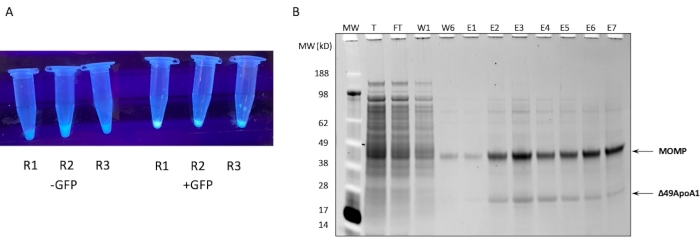
Figure 1: Expression and purification of MOMP-tNLP. (A) Image of tubes containing small aliquots of a cell-free reaction that successfully expressed GFP controls luminating under UV light source (right) compared to lysates without GFP plasmid (left). (B) Protein Gel stained with SYPRO Ruby after SDS-PAGE shows the purification profile of MOMP-tNLP. MOMP migrates at 40 kDa and the 49ApoA1 migrates at 22 kDa. Abbreviations: MOMP = chlamydial major outer membrane protein; tNLP = telodendrimer nanolipoprotein particle; MOMP-tNLP = MOMP-tNLP complex; GFP = green fluorescent protein-encoding plasmid; MW = Molecular weight marker; T = total cell-free lysate; FT = flow-through; R1-R3 = cell-free reaction aliquots; W1, W6 = Washes 1 and 6; E1-E7 = Elutions 1 through 7; Δ49ApoA1 = His-tagged mouse ApoA1 derivative. Please click here to view a larger version of this figure.
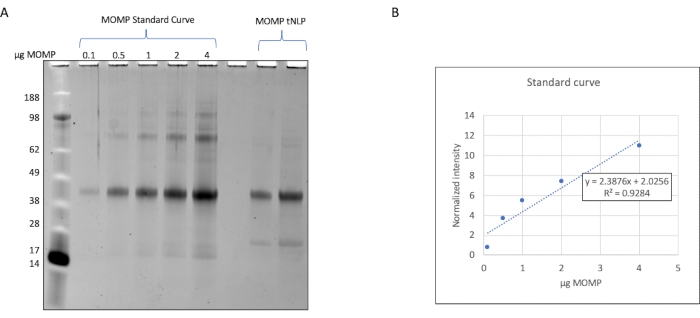
Figure 2: Quantification of MOMP in MOMP-tNLP samples. (A) SDS-PAGE gel stained with SYPRO Ruby for the quantification of MOMP. Recombinant MOMP with known concentration was loaded onto the gel to obtain the standard curve. Each lane contained 0.1 µg, 0.5 µg, 1.0 µg, 2.0 µg, and 4.0 µg of MOMP. MOMP-tNLP samples that were being quantified were loaded on the same gel. (B) The MOMP concentration standard curve was generated using densitometry. An equation relating normalized band density and the amount of MOMP was established. The equation was used to calculate the MOMP content in the unknown samples. Abbreviations: MOMP = chlamydial major outer membrane protein; tNLP = telodendrimer nanolipoprotein particle; MOMP-tNLP = MOMP-tNLP complex; SDS-PAGE = sodium dodecylsulfate polyacrylamide gel electrophoresis. Please click here to view a larger version of this figure.
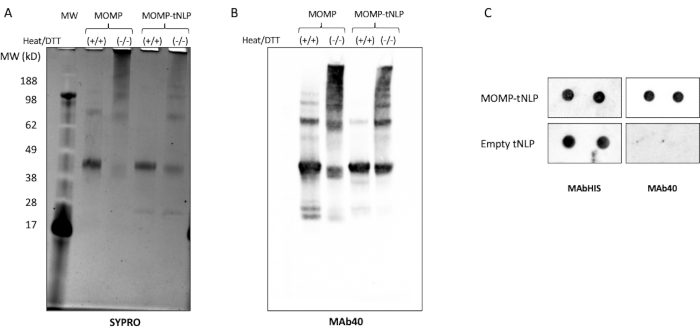
Figure 3: Cell-free-produced MOMP-tNLP allow MOMP to form higher-order structures. (A) SDS-PAGE gel of MOMP-tNLP with and without treatment of heat and reducing agent DTT, stained with SYPRO Ruby. With heat and DTT, MOMP primarily appeared as a monomer band at ~40 kDa, as heat and the reducing agent broke down the majority of higher-order MOMP structure. In the absence of heat and DTT, the higher-order bands were present, indicating MOMP oligomer conformation. (B) Western blot of MOMP-tNLP and MOMP alone, untreated and treated with heat and DTT. After transfer, the membrane was probed with MAb40 (1:1,000 dilution). A banding pattern similar to the SYPRO Ruby-stained gel was observed, confirming that the higher molecular weight bands were indeed MOMP oligomers. (C) Dot blot of MOMP-tNLP and empty tNLP samples (in duplicate) probed with MAb40 and MAbHIS. Abbreviations: MOMP = chlamydial major outer membrane protein; tNLP = telodendrimer nanolipoprotein particle; MOMP-tNLP = MOMP-tNLP complex; SDS-PAGE = sodium dodecylsulfate polyacrylamide gel electrophoresis; DTT = dithiothreitol. Please click here to view a larger version of this figure.
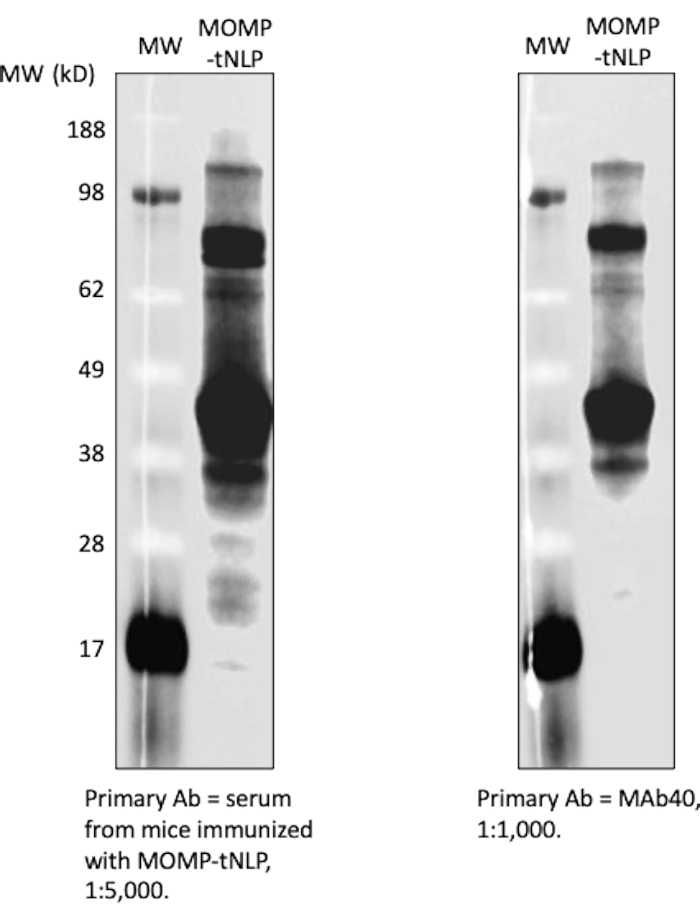
Figure 4: Cell-free-produced MOMP-tNLP is highly immunogenic. Serum from immunized mice showed strong anti-MOMP IgG signal. MOMP-tNLP adjuvanted with CpG + FSL-1 was used to immunize mice. Sera from six immunized mice were collected, pooled, and used to probe MOMP-tNLP. The serum was able to bind to MOMP in a western blotting assay and showed strong IgG signal (left). The western blot using MAb40 as primary antibody (right) showed similar bands, indicating that the serum contained MOMP-specific IgG. Abbreviations: MOMP = chlamydial major outer membrane protein; tNLP = telodendrimer nanolipoprotein particle; MOMP-tNLP = MOMP-tNLP complex; CpG = cholesterol-modified CpG adjuvant; FSL-1 = lipophilic adjuvant. Please click here to view a larger version of this figure.
| Buffer name | NaH2PO4 | NaCl | Imidazole | pH |
| Binding Buffer | 50 mM | 300 mM | 10 mM | 8.0 |
| Wash Buffer | 50 mM | 300 mM | 20 mM | 8.0 |
| Elution Buffer 1 | 50 mM | 300 mM | 250 mM | 8.0 |
| Elution Buffer 2 | 50 mM | 300 mM | 500 mM | 8.0 |
Table 1: List of buffers needed for nickel affinity purification detailing concentrations of each component and pH.
| Runtime | 50 min |
| Flow rate | 6.0 mL/min |
| Gradient type | Binary |
| Buffer A | 10 mM TEAA in H20 |
| Buffer B | MeCN |
| Gradient | % Buffer B |
| 0 min | 25% |
| 30 min | 60% |
| 30.5 min | 100% |
| 40 min | 100% |
| 40.5 min | 25% |
| 50 min | 25% |
Table 2: Conditions for reversed phase HPLC purification of cholesterol-modified CpG. Abbreviations: TEAA = triethylammonium acetate; MeCN = acetonitrile.
| Cell-free lysate (mL) | DMPC lipid (mg) | Telodendrimer (mg) | MOMP plasmid (µg) | Purified MOMP yield (mg) |
| 1 | 4 | 0.4 | 15 | 0.5 |
| 2 | 8 | 0.8 | 30 | 1.1 |
| 3 | 12 | 1.2 | 45 | 1.6 |
| 5 | 20 | 2 | 75 | 2.7 |
Table 3: The quantity of lipids, telodendrimer, and plasmids used for differently scaled cell-free reactions and the corresponding yields. Abbreviations: MOMP = chlamydial major outer membrane protein; DMPC = 1,2-dimyristoyl-sn-glycero-3-phosphocholine.
| Ratios of plasmid input, MOMP : Δ49ApoA1 | 1:1 | 5:1 | 10:1 | 25:1 | 50:1 | 100:1 | |
| Ratios of the amount of protein produced, MOMP : Δ49ApoA1 | 0.02 | 0.32 | 0.64 | 3.46 | 6.55 | 20.04 | |
| Estimated number of MOMP insertion per tNLP | 0.03 | 0.37 | 0.75 | 4.04 | 7.65 | 23.39 | |
Table 4: The plasmid ratios in a cell-free reaction and the resulting MOMP insertion rates. Abbreviations: MOMP = chlamydial major outer membrane protein; tNLP = telodendrimer nanolipoprotein particle; Δ49ApoA1 = His-tagged mouse ApoA1 derivative.
