Analysis of Cell Differentiation, Morphogenesis, and Patterning During Chicken Embryogenesis Using the Soaked-Bead Assay
Summary
The soaked bead assay involves targeted delivery of test reagent at any developmental time point to study the regulation of cell differentiation and morphogenesis. A detailed protocol, applicable to any experimental animal model, for preparing three different types of soaked beads and implanting these in the interdigit of a chicken embryo is presented.
Abstract
A multitude of genetic programs is activated during embryonic development that orchestrates cell differentiation to generate an astounding diversity of somatic cells, tissues, and organs. The precise activation of these genetic programs is regulated by morphogens, diffusible molecules that direct cell fate at different thresholds. Understanding how genetic activation coordinates morphogenesis requires the study of local interactions triggered by morphogens during development. The use of beads soaked in proteins or drugs implanted into distinct regions of the embryo enables studying the role of specific molecules in the establishment of digits and other developmental processes. This experimental technique provides information on the control of cell induction, cell fate, and pattern formation. Thus, this soaked bead assay is an extremely powerful and valuable experimental tool applicable to other embryonic models.
Introduction
Breakthroughs in the molecular mechanisms that control gene expression during embryonic development have allowed us to understand how cell fate is determined. Commitment to different cell lineages occurs once cells begin the molecular expression of transcription factors1. This expression pattern is highly coordinated in space and time and thereby directs the shaping, positioning, and patterning of cells, tissues, and organs1,2,3,4,5. Embryonic induction is the process by which cells are committed to specific lineages by establishing hierarchies that restrict cells' potentiality, which even include the generation of the basic body plan as occurs with the Spemann organizer6,7. The blastopore dorsal lip induces a second embryonic axis in a host embryo8,9. Today, with the aid of grafting and other classical experiments combined with molecular approaches, it is known that different transcription factors and growth factors function to direct embryonic induction in the Spemann organizer10. Thus, experimental manipulation is an important tool to understand cell differentiation, morphogenesis, and patterning processes during embryogenesis.
Interestingly, in embryonic systems where tissue transplantation is difficult or when the inducers are already well known, carriers are used to deliver molecules (e.g., proteins, chemicals, toxins, etc.) to regulate cell differentiation, morphogenesis, and even patterning. One such carrier system involves implanting beads soaked in a specific molecule in any experimental model organism at any developmental time point to determine the effect of the said reagent or direct the differentiation of the said model. For example, by implanting retinoic acid (RA)-soaked beads into the chicken wing limb bud, Cheryl Tickle et al. (1985) demonstrated that RA induces the expression of sonic hedgehog in the zone of polarizing activity (ZPA)11,12. The same experimental strategy was used to discover that RA controls the asymmetry of somites and cell death in the limb bud during digit development and in other embryonic limb regions13,14,15. Other factors, mainly proteins (e.g., fibroblast growth factors [FGF], transforming growth factor-beta [TGF-ß]) have been used to induce limbs in early embryos' flanks and new digits in the interdigital region, respectively16,17,18,19,20,21. These experiments evidence the power and utility of this technique for determining the stage of commitment or competence of tissues or groups of cells exposed to the molecules.
In this protocol, the chick limb at the stage of digit formation served as the experimental model to present step-by-step how to prepare and implant the soaked beads. However, this experimental tool is not limited to this application but can be exploited in any experimental animal model and any timepoint in vitro and in vivo to study induction, differentiation, cell death, and patterning.
Protocol
This research was reviewed and approved by the Institutional Review Board for the Care and Use of Laboratory Animals of the Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México (UNAM, Mexico City, Mexico).
1. Egg incubation and embryo staging
NOTE: Fertilized hen eggs can be obtained from local farms. Fertilized White Leghorn chicken eggs are most commonly used. Store the freshly fertilized chicken eggs at 15 °C for up to 1 week prior to incubation.
- Incubate the fertilized chicken eggs vertically with the pointy side down in a humidified incubator at 38 °C and 70% relative humidity until they reach the 28 HH stage (approximately 5.5 days according to Hamilton and Hamburger, 1951)22. Rotate the eggs during the incubation to prevent the embryo from adhering to the shell membrane.
NOTE: The choice of the developmental stage is informed by the experimental aims. In this case, the 28 HH stage is optimal for interdigit bead implantation to induce an ectopic digit or promote cell death. - Remove the eggs from the incubator, swab them with 70% ethanol, and allow them to air dry. Disinfect the working area, microscopes, and instruments with 70% ethanol.
- Candle the egg to identify blood vessels and locate the embryo. Discard eggs that do not have an embryo.
- Using the end of non-toothed forceps, open a window in around five eggs by tapping the blunt end of an egg, and remove a 1-cm2 section of the shell with the forceps.
- Transfer the egg into a carton or plastic holder and place it under the stereomicroscope. Remove the air membrane by puncturing and pulling it out with fine surgical forceps. Remove any small piece of eggshell that could contact the embryo.
NOTE: The air membrane is the white, opaque membrane observed immediately after windowing the egg. - Observe under the microscope while opening the amniotic sac slightly, tearing it using the fine surgical forceps. Be careful not to damage the vasculature of the chorioallantoic membrane.
NOTE: The amnion is the transparent membrane closely surrounding the embryo that encapsulates it in amniotic fluid. - Stage the embryos in ovo to determine whether they are in the desired stage. Embryos in earlier stages can be returned to the incubator after sealing the eggshell window with tape.
2. Bead preparation
NOTE: Depending on the experimental aim and treatment in question, alternative bead types (e.g., Affi-Gel, AG1-X2, heparin) may be more suitable. Affi-Gel beads are optimal for proteins (e.g., TGF-ß1), while heparin beads are ideal for growth factors (e.g., FGFs, WNT) and AG1-X2 beads for chemicals solubilized in organic solvents (e.g., DMSO).
- Affi-Gel and heparin bead preparation
- Cut a square of parafilm to fit a 45-mm Petri dish. Place the parafilm across the bottom of the petri dish to cover it and fix it to the bottom of the Petri dish by pushing each vertex using the end of non-toothed forceps. Set aside.
- Using a pipette or spatula, transfer the beads into a microcentrifuge tube and wash them twice in 1x PBS by settling and pipetting.
- Transfer ~40-50 beads with a micropipette to the center of the parafilmed Petri dish from step 1.
- Using the microscope, select ~30 Affi-Gel or heparin beads ~100 µm in diameter for use. Use a microscope eyepiece reticle to size the beads or use the third interdigit of an HH 28 embryo as a reference; the bead must be smaller than the interdigit.
- Carefully remove as much of the excess PBS surrounding the beads as possible and soak them in 2-5 µL of the treatment solution. Assure that the solution completely covers beads.
- In parallel, prepare control beads by soaking in a solution that contains the same amount of vehicle as used in the experimental treatment solution.
NOTE: Concentrations need to be calculated for each treatment according to the experiment. Use the appropriate personal protective equipment when handling potentially harmful reagents. - Incubate the beads in the solution for 30 min at room temperature. To prevent the beads from drying out during the incubation, pipette a few drops of 1x PBS or water around the beads to humidify the local atmosphere and cover the dish with parafilm to slow evaporation.
- Place the Petri dish on ice and implant the beads within the same day.
- AG 1-X2 bead preparation
- Cut a square of parafilm to fit a 45-mm Petri dish. Cover the bottom of the Petri dish with parafilm and affix by pushing each vertex using the end of non-toothed forceps. Set aside.
- Use a spatula to transfer the AG 1-X2 beads into a microcentrifuge tube. Add 30-50 µL of the treatment solution at the desired final concentration.
- In parallel, incubate the control beads in a solution with the vehicle alone, prepared without the experimental chemical or protein of interest.
- Incubate the beads for 20 min while slowly shaking at room temperature. Wrap the microcentrifuge tubes with foil to protect from light during all incubation, given that many of these molecules are light-sensitive
- Remove as much of the solution as possible using a pipette and stain with 2% phenol red dissolved in water at room temperature for 2 min with mild agitation in a vortex.
NOTE: Dyeing the transparent AG 1-X2 beads facilitates their implantation in the embryo. - Remove the 2% phenol red and wash the beads twice in 1x PBS to remove any excess dye.
- With a micropipette tip, transfer ~40-50 AG 1-X2 beads to the center of the parafilm-covered Petri dish prepared in step 1 and soak these in 5 µL of 1x PBS. Under the microscope, select around 30 beads that are ~100 µm diameter in size. Discard the unused beads.
- Beads are ready to implant. Ensure the beads are submerged in the PBS throughout the implantation by pipetting a few drops of 1x PBS or water around the beads to humidify the local atmosphere and/or cover the dish with parafilm to slow evaporation.
3. Embryo manipulation and interdigit implantation
- Before manipulating the embryos, arrange two stereomicroscopes next to each other on a benchtop. One is for embryo manipulation and bead implantation, and the other is for maintaining the treated beads ready to implant into the embryo.
- Using non-toothed forceps, create a window in the remaining eggs as described in the egg incubation and embryo staging section.
- Under the microscope, open the amniotic sac by tearing the amniotic membrane with fine surgical forceps near the right hindlimb only by the amount needed to accomplish the procedure (i.e., as little as possible).
- Using fine forceps, hold the embryo by the amniotic membrane to expose the right hindlimb. Using a fine tungsten needle, make a hole centered in the distal-most of the third interdigit of the hindlimb.
- Without releasing the embryo and the exposed hindlimb, take one treated bead from the other microscope using one of the tips of the forceps. The forceps must be in the open position for one bead to adhere to the forceps tip.
- Transfer the bead into the chick embryo near the hindlimb and position it on top of the interdigit hole.
- Close the forceps to apply pressure to the bead until it enters the hole.
- Release the embryo and seal the eggshell window with tape.
- Return the eggs to the incubator until they have reached the required stage. Repeat the procedure until the required number of manipulated embryos is reached. It is imperative to ensure that the beads do not dry out at any point.
NOTE: To observe a complete ectopic finger, incubate for ~72 h after bead implantation. In contrast, early differentiation genes (e.g., Sox9) are expressed ~30 min after the implantation of a TGF-ß1-soaked bead.
Representative Results
Using soaked beads to evaluate cell behavior in the embryonic chick limb
To assure the efficacy of this assay, the bead must be placed consistently and precisely in the correct location; in this case, the distal-most of the third interdigit beneath the apical ectodermal ridge AER (Figure 1A). This positioning permits the molecule in question to spread equally throughout the interdigital tissue. Moreover, the zone beneath the AER contains undifferentiated cells that are readily responsive to treatment. To evaluate the effects on cell differentiation, the embryos can be stained with Alcian blue and Alizarin red to evidence the formation of skeletal elements (Figure 1B). The soaked-bead assay is also well-suited for evaluating cell death with neutral red (Figure 1C) and gene regulation by in situ hybridization (Figure 1D). Scale bar is set at 250 µm.
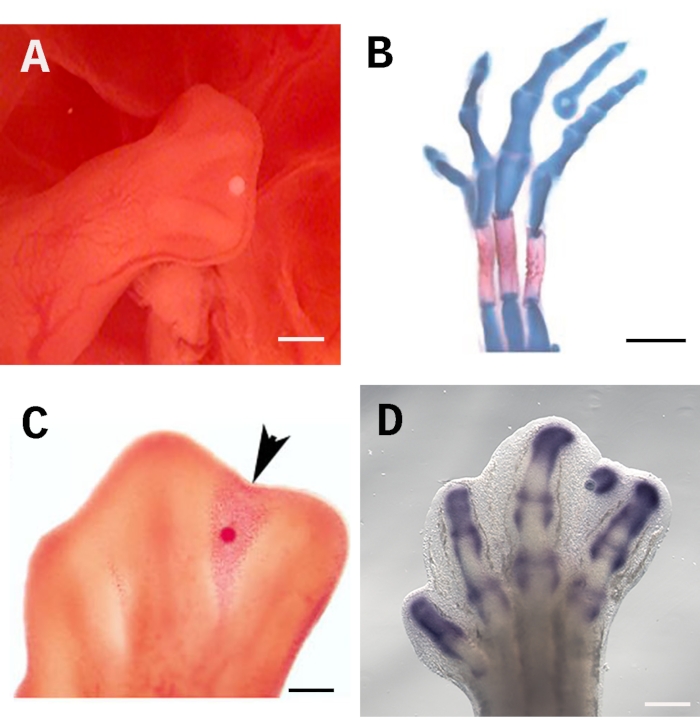
Figure 1: Bead implantation into the interdigital tissue of the chick limb.(A)The correct location for the soaked bead beneath the AER in a 28 HH hindlimb. (B) Alcian blue and Alizarin red skeletal staining evidence the formation of an ectopic digit induced 4 days after implanting a TGF-ß1-soaked bead. (C) Neutral red staining marks cell death induction (arrowhead) 24 h after implanting a RA-soaked bead into the interdigital tissue at the 28 HH stage. (D) Sox9 in situ hybridization 4 h after a TGF-ß1-soaked bead was implanted. Scale bar is set at 250 µm. The images shown in B and C were taken from Díaz-Hernández et al.23,24. Please click here to view a larger version of this figure.
Discussion
The main advantage of the experimental tool detailed in this protocol is being able to control the time and location of the exposure to beads soaked in a given experimental molecule. Combining the correct positioning with precise developmental timing provides enormous possibilities to study cell differentiation processes. Performing these experiments in undifferentiated tissue enables investigating the first crucial events in cellular lineage. For example, placing a TGFß-soaked bead in the interdigital tissue of embryonic limbs 28 HH results in the formation of an ectopic digit in which a molecular cascade is triggered that induces the genetic expression of the master gen Sox925. Remarkably, the induced cartilage tissue also organizes into a digit with phalanx formation.
Interestingly, RA triggers cell death in the same interdigital region by regulating the gene expression of bone morphogenetic proteins that direct the cell fate of undifferentiated cells toward cell death10. Hence, cell differentiation, cell death, morphogenesis, and patterning can be concurrently investigated in the same region of an embryo and tailored to any region and genetic pathway of interest8,9.
The elements crucial to the success of this protocol include never letting the soaked beads dry out (i.e., they must always remain wet). Also, selecting the appropriate beads is essential: Affi-Gel and heparin beads are for proteins, whereas AG1-X2 are for chemicals dissolved in organic solvents. Another critical point is the concentration of the molecule contained in the solution used to soak the beads, which is usually 1000-fold more concentrated than would be used for in vitro studies. Nevertheless, an inconvenience of this method is that the final concentration of molecules released from the soaked beads is unknown, as well as the velocity of release. In the protocol is mentioned that beads-100 µm diameter is more convenient for use. Consider this when limbs are manipulated. Most importantly, the diameter of the beads must be selected according to the implantation zone and consistently maintained in each embryo. However, a slight variation in bead size between experiments is not likely to affect the results.
In conclusion, the potential of the soaked-bead assay outlined here depends only on the imagination of the researcher. This protocol can be applied to any experimental animal, cell culture, or organotypic culture model, including organoids. Furthermore, this protocol is a helpful, straightforward educational tool for teaching students basic developmental biology concepts and technical skills by practicing this experimental manipulation in developmental biology classes.
Offenlegungen
The authors have nothing to disclose.
Acknowledgements
This work was supported by the Dirección General de Asuntos del Personal Académico (DGAPA)-Universidad Nacional Autónoma de México [grant numbers IN211117 and IN213314] and Consejo Nacional de Ciencia y Tecnología (CONACyT) [grant number 1887 CONACyT-Fronteras de la Ciencia] awarded to JC-M. JC M-L received a postdoctoral fellowship from the Consejo Nacional de Ciencia y Tecnología (CONACyT-Fronteras de la Ciencia-1887). The authors appreciate the help of Lic. Lucia Brito from Instituto de Investigaciones Biomédicas, UNAM in the preparation references of this manuscript.
Materials
| Affi-Gel Blue Gel beads | Bio-Rad | 153-7302 | |
| AG1-X2 beads | Bio-Rad | 1400123 | |
| Egg incubator | Incumatic de Mexico | Incumatic 1000 | |
| Fine surgical forceps | Fine Science Tools | 9115-10 | |
| Heparine Sepharose beads | Abcam | ab193268 | |
| Petri dish | Nest | 705001 | |
| Stereomicroscope | Zeiss | Stemi DV4 | |
| Tape | NA | NA | |
| Tungsten needle | GoodFellow | E74-15096/01 |
Referenzen
- Stapornwongkul, K. S., Vincent, J. P. Generation of extracellular morphogen gradients: the case for diffusion. Nature Reviews Genetics. 22 (6), 393-411 (2021).
- Rogers, K. W., Schier, A. F. Morphogen gradients: from generation to interpretation. Annual Review of Cell and Developmental Biology. 27, 377-407 (2011).
- Irizarry, J., Stathopoulos, A. Dynamic patterning by morphogens illuminated by cis-regulatory studies. Development. 148 (2), 196113 (2021).
- Capek, D., Müller, P. Positional information and tissue scaling during development and regeneration. Development. 146 (24), (2019).
- Marín-Llera, J. C., Garciadiego-Cázares, D., Chimal-Monroy, J. Understanding the cellular and molecular mechanisms that control early cell fate decisions during appendicular skeletogenesis. Frontiers in Genetics. 10, 977 (2019).
- Gurdon, J. B. Embryonic induction–molecular prospects. Development. 99 (3), 285-306 (1987).
- Bouwmeester, T. The Spemann-Mangold organizer: the control of fate specification and morphogenetic rearrangements during gastrulation in Xenopus. International Journal of Developmental Biology. 45 (1), 251-258 (2001).
- Piccolo, S., Sasai, Y., Lu, B., De Robertis, E. M. Dorsoventral patterning in Xenopus: inhibition of ventral signals by direct binding of chordin to BMP-4. Cell. 86 (4), 589-598 (1996).
- Cho, K. W., Blumberg, B., Steinbeisser, H., De Robertis, E. M. Molecular nature of Spemann’s organizer: the role of the Xenopus homeobox gene goosecoid. Cell. 67 (6), 1111-1120 (1991).
- Thisse, B., Thisse, C. Formation of the vertebrate embryo: Moving beyond the Spemann organizer. Seminars in Cell & Development Biology. 42, 94-102 (2015).
- Eichele, G., Tickle, C., Alberts, B. M. Microcontrolled release of biologically active compounds in chick embryos: beads of 200-microns diameter for the local release of retinoids. Analytical Biochemistry. 142 (2), 542-555 (1984).
- Tickle, C., Lee, J., Eichele, G. A quantitative analysis of the effect of all-trans-retinoic acid on the pattern of chick wing development. Entwicklungsbiologie. 109 (1), 82-95 (1985).
- Vermot, J., Pourquié, O. Retinoic acid coordinates somitogenesis and left-right patterning in vertebrate embryos. Nature. 435 (7039), 215-220 (2005).
- Rodriguez-Leon, J., et al. Retinoic acid regulates programmed cell death through BMP signalling. Nature Cell Biology. 1 (2), 125-126 (1999).
- Rodriguez-Guzman, M., et al. Tendon-muscle crosstalk controls muscle bellies morphogenesis, which is mediated by cell death and retinoic acid signaling. Entwicklungsbiologie. 302 (1), 267-280 (2007).
- Cohn, M. J., Izpisúa-Belmonte, J. C., Abud, H., Heath, J. K., Tickle, C. Fibroblast growth factors induce additional limb development from the flank of chick embryos. Cell. 80 (5), 739-746 (1995).
- Ohuchi, H., et al. An additional limb can be induced from the flank of the chick embryo by FGF4. Biochemical and Biophysical Research Communications. 209 (3), 809-816 (1995).
- Abu-Elmagd, M., Goljanek Whysall, K., Wheeler, G., Münsterberg, A. Sprouty2 mediated tuning of signalling is essential for somite myogenesis. BMC Medical Genomics. 8, 8 (2015).
- Gañan, Y., Macias, D., Duterque-Coquillaud, M., Ros, M. A., Hurle, J. M. Role of TGF beta s and BMPs as signals controlling the position of the digits and the areas of interdigital cell death in the developing chick limb autopod. Development. 122 (8), 2349-2357 (1996).
- Merino, R., et al. Morphogenesis of digits in the avian limb is controlled by FGFs, TGFbetas, and noggin through BMP signaling. Entwicklungsbiologie. 200 (1), 35-45 (1998).
- Montero, J. A., Lorda-Diez, C. I., Gañan, Y., Macias, D., Hurle, J. M. Activin/TGFbeta and BMP crosstalk determines digit chondrogenesis. Entwicklungsbiologie. 321 (2), 343-356 (2008).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88 (1), 49-92 (1951).
- Díaz-Hernández, M. E., Bustamante, M., Galván-Hernández, C. I., Chimal-Monroy, J. Irx1 and Irx2 are coordinately expressed and regulated by retinoic acid, TGFβ and FGF signaling during chick hindlimb development. PLoS One. 8 (3), 58549 (2013).
- Díaz-Hernández, M. E., Rios-Flores, A. J., Abarca-Buis, R. F., Bustamante, M., Chimal-Monroy, J. Molecular control of interdigital cell death and cell differentiation by retinoic acid during digit development. Journal of Developmental Biology. 2 (2), 138-157 (2014).
- Chimal-Monroy, J., et al. Analysis of the molecular cascade responsible for mesodermal limb chondrogenesis: Sox genes and BMP signaling. Entwicklungsbiologie. 257 (2), 292-301 (2003).
