Morphology of the cultured cells was observed from the start of the culture until BOEC colonies were observed (Figure 1). A smaller population of adherent cells started to attach to the culture dishes and grow, while non-adherent cells were removed with culture medium changes (Figure 1B). Colonies first appeared on day 6 as a collection of endothelial-like cells proliferating radially outward from a central point (Figure 1D). As the culture progressed, cell colonies became more dense and displayed a cobblestone morphology similar to mature endothelial cells (Figure 1F). The typical endothelial cobblestone morphology provides an easy preliminary identification of BOECs using a light microscope.
Endothelial cell colonies are typically ready for passaging at 10-14 days of culture. At this time, the 6-well plate will typically yield ~1 million cells with percent viability of 93%-98%. During the first passage, endothelial cells will expand to yield ~6-10 million cells in a T75 flask.
To assess the morphogenesis of BOECs, the basement membrane matrix (e.g., Matrigel) was plated on 15 well angiogenesis plates at 10 µL/well and incubated at 37 °C for 30 min to allow polymerization. Passage 2 cells were seeded on basement membrane matrix-coated plates and imaged at 14 h and 24 h time points. A density of approximately 3,500 cells per well was able to form a network and tube-like structures. Tube formation was noticed within 14 h for serum free medium (EGM-2 with supplementals except for FBS) and within 24 h for complete growth medium (EGM-2 with supplements including FBS). Adding FBS may have diluted the endothelial cell-specific growth factors (e.g., VEGF) and introduced other signaling factors resulting in the delay of the tube formation process. 2D phase-contrast microscopy was used to image tube formation in serum-free medium (Figure 2A,B) and complete growth medium (Figure 2C,D). Cells in both media conditions underwent morphological differentiation and rapidly organized into extensive networks of capillary tube-like structures. These structures were composed of organized cellular cords resembling in vivo capillary networks. Furthermore, micrographs at 20x magnification illustrate the complex multicellular organization of endothelial cells and their morphological differentiation in detail. There were no morphological differences observed between the media conditions. The extensive network of capillary tube-like structures strongly suggests the differentiation of cultured cells into mature and functional endothelial cells.
BOECs were further characterized by the expression of mature endothelial cell marker CD31 or platelet endothelial cell adhesion molecule-1 (PECAM1) (Figure 3A,B). BOECs showed uniform expression of CD31. Furthermore, flow cytometry analysis confirmed the absence of EPCs as the cells stained negative for monocyte marker CD14 compared to the positive control group of PBMCs (Figure 3C,D).
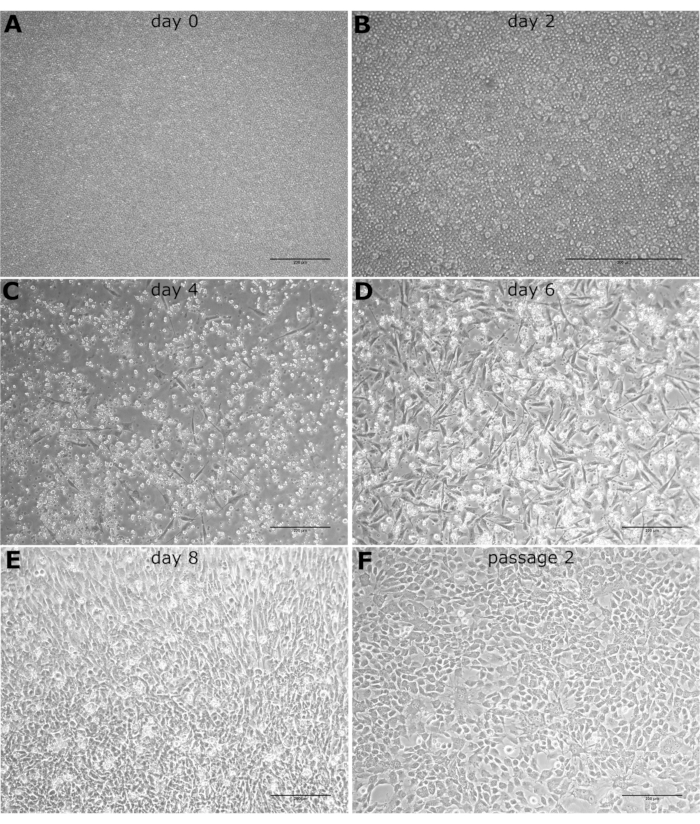
Figure 1: Phenotype light microscopy images. The light microscopy images of cultured BOECs observed at 10x magnification on (A) day 0, (B) day 2, (C) day 4, (D) day 6, (E) day 8, and (F) after the first passage. Scale bars: 200 µm. Please click here to view a larger version of this figure.
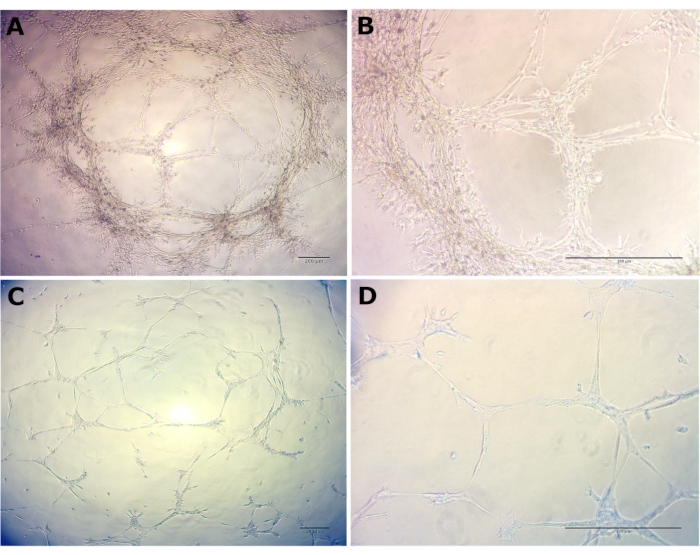
Figure 2: Morphological differentiation and organization of passage 2 BOECs. Morphological differentiation and organization of passage 2 BOECs into capillary tube-like structures was noticed in the basement membrane matrix within 14 h for serum-free medium and within 24 h for complete growth medium. (A) Serum-free medium at 4x, (B) Serum-free medium at 20x, (C) Complete growth medium at 4x, and (D) Complete growth medium at 20x. Scale bars: 200 µm. Please click here to view a larger version of this figure.
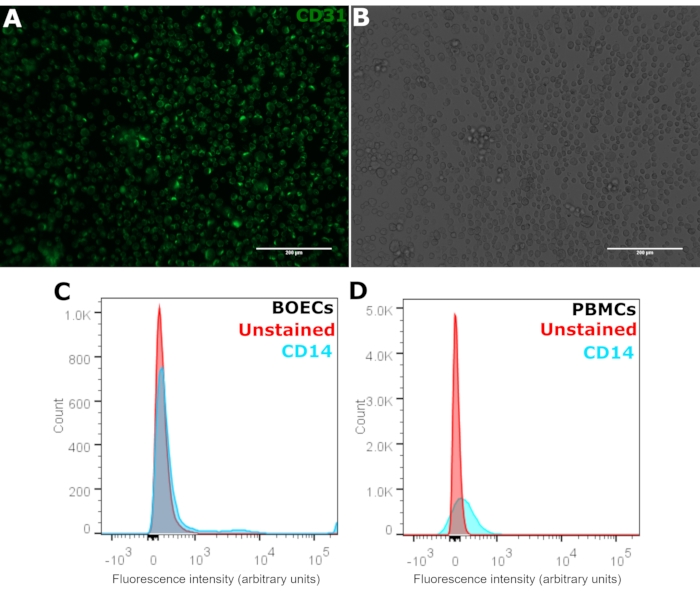
Figure 3: Characterization of BOECs with CD31 and CD14. (A) Passage 2-3 BOECs were stained for PECAM1 using anti-CD31-FITC antibody seen in green and (B) corresponding phase-contrast microscopy images. Cells were stained in suspension, and the suspension was imaged on a microscope slide. Scale bars: 200 µm. (C) Flow cytometry analysis of BOECs with CD14-AF700 antibody compared to (D) positive control peripheral blood mononuclear cells (PBMCs). Positive CD14 staining shown in blue and unstained cells shown in red. Please click here to view a larger version of this figure.
