The protocol used here is summarized in the flowchart in Figure 1. The inhalation of clodronate liposomes through the nose (Figure 2) was used to deplete the pulmonary macrophages of adult C57BL/6 mice, and this produced a good recipient mouse model. Pulmonary IMs were isolated from another untreated (host) mouse (Figure 3A,B) and cultured in vitro. The isolated macrophages were stimulated with IL-33 for 24 h and then intratracheally instilled into the recipient mice (Figure 3C), with unstimulated macrophages used as a control. After 24 h, BLM was administered to the recipient mice to induce a pulmonary fibrosis model. The extent of pulmonary fibrosis after the adoptive transfer of IMs with or without IL-33 stimulation was compared 21 days after BLM administration. Hematoxylin and eosin (H&E) staining of the pathological tissue sections showed that the typical pathological changes of fibrosis were observed after BLM administration: the lung tissue structure of the mice was destroyed, the aggregation of fibroblasts was observed, and the normal alveoli disappeared or decreased. The adoptive transfer of IL-33-stimulated macrophages exacerbated the degree of lung tissue destruction and increased fibroblast aggregation in the BLM-stimulated mice (Figure 4A). The increase in the Ashcroft score further illustrated the degree of pulmonary fibrosis (Figure 4B). The pathological process of pulmonary fibrosis is associated with the increased aggregation of myofibroblasts, which secrete α-smooth muscle actin (α-SMA) and fibronectin. The determination of the expression levels of these markers showed that the mRNA levels of α-SMA (Figure 5B) and fibronectin (Figure 5A) in the lung tissues of the recipient mice containing adoptively transferred IL-33-stimulated IMs and treated with BLM were further increased compared with those of the BLM-treated wild-type mice.
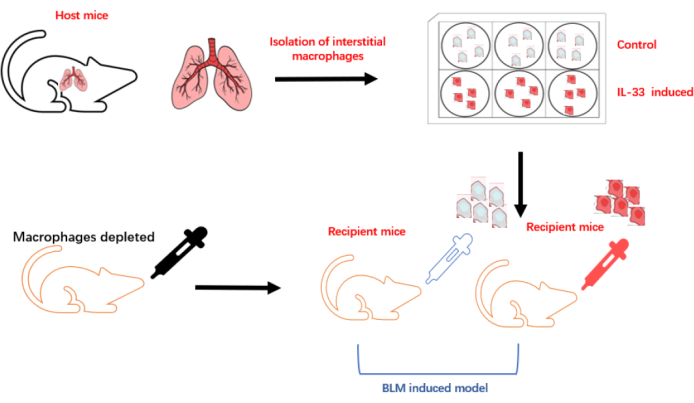
Figure 1: Schematic diagram of the establishment of the macrophage adoptive transfer experiment. Please click here to view a larger version of this figure.
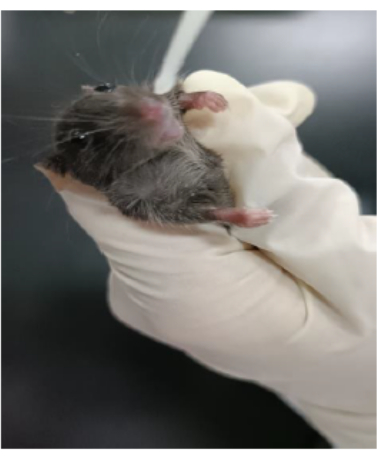
Figure 2: Depletion of macrophages in the recipient mice. The clodronate liposomes were dropped into the nasal cavity of the mice. Please click here to view a larger version of this figure.
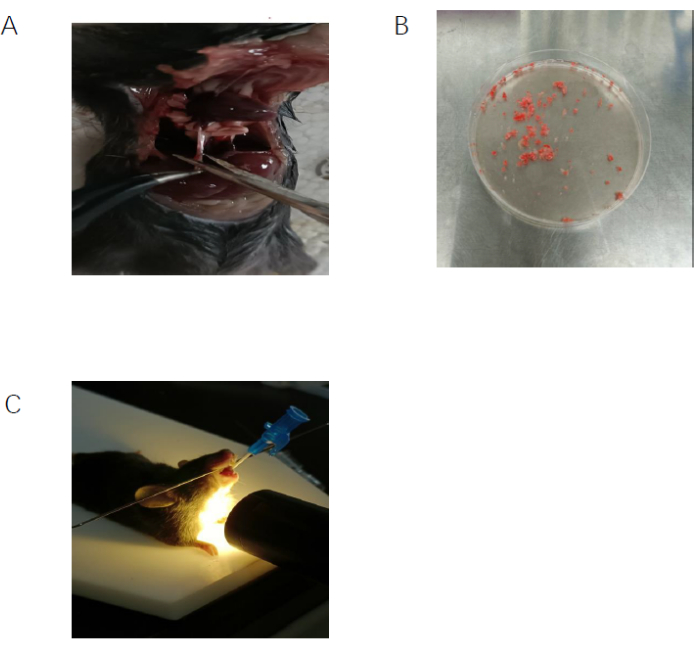
Figure 3: Isolation and adoptive transfer of interstitial macrophages. (A) The inferior vena cava of the host mouse was cut to facilitate perfusion. (B) The lung tissue from the host mouse was cut into small pieces. (C) The interstitial macrophages were purified to 94% using flow cytometry with F4/80 and CD11c markers. (D) IL-33-stimulated IMs were administered to the recipient mice through the trachea. Please click here to view a larger version of this figure.
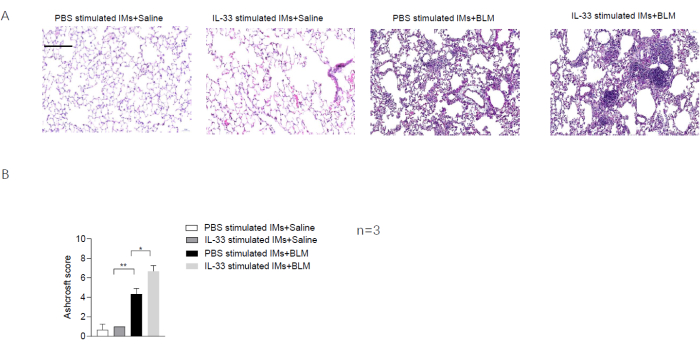
Figure 4: The effect of the adoptive transfer of IL-33-stimulated macrophages in the BLM-stimulated mice. (A) H&E staining of lung tissue sections from the recipient mice. Scale bar = 100 µm. (B) Ashcroft scores determined from the lung tissue sections of the recipient mice. Data are shown as mean ± SEM (n = 3). *p < 0.05, **p < 0.01. Please click here to view a larger version of this figure.
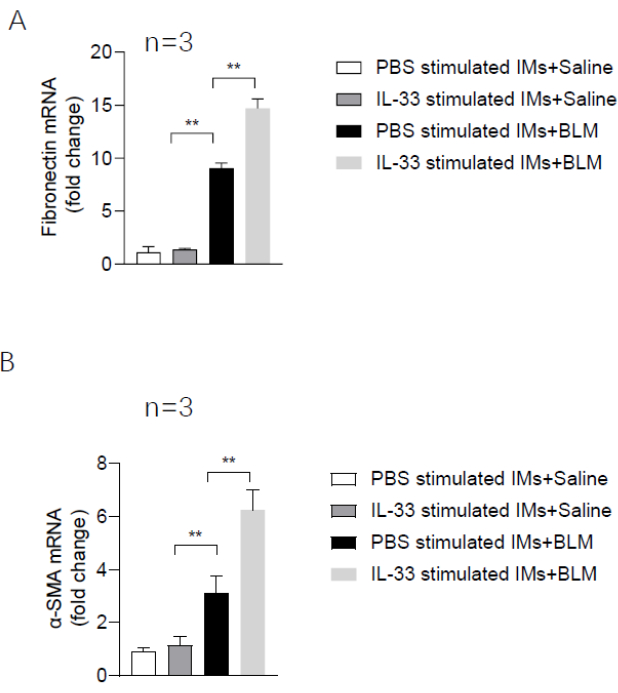
Figure 5: Expression levels of pulmonary fibrosis marker genes in the recipient mice. (A) The mRNA level of fibronectin. (B) The mRNA level of α-SMA. Data are shown as mean ± SEM (n = 3). *p < 0.05, **p < 0.01. Please click here to view a larger version of this figure.
Supplementary Figure 1: Depletion of alveolar macrophages and purity of IMs. (A) The depletion of alveolar macrophages was confirmed by checking the expression of F4/80 and CD11b markers. (B) The purity of the obtained IMs was assessed using flow cytometry with staining for F4/80 and CD11c markers, and the purity of the isolated IMs was determined to be 94.4% using flow cytometry. Please click here to download this File.
