Selecting the embryo maturation schedule for the transplant
The experiment begins at D0 when fertilized eggs are incubated at 38 °C and 60% relative humidity. The chorioallantoic membrane (CAM) is a highly vascularized extraembryonic membrane that develops after egg incubation. It is formed by the fusion of the allantois and chorion. At D1, after 24 h of incubation, the air chamber is punctured to prevent the CAM from attaching to the inner shell membrane. Puncturing the air chamber at D1 improves the quality of the air chamber compared to puncture at later stages (D4). The CAM continues to grow until D12 when it envelops the entire egg content and adheres firmly to the inner shell membrane (Figure 1). The hole made at D7 is enlarged to facilitate sample harvest. D7 is the optimal time for grafting when the CAM reaches maturity and covers the yolk sac and the embryo surface. At this stage, the air chamber hole is enlarged, and the organoids are transferred into the CAM with the help of an automatic pipette.
Brain organoid grafting
Throughout the inherent maturation of brain organoids in vitro, their cellular composition changes from neural progenitors to mature neurons and glial cells. Neurovasculature and neurogenesis are intermingled and interdependent developmental processes in vertebrates, and the signaling leading to this crosstalk is highly dependent on the maturation stage of the neural counterpart29. Neural progenitors are the major source of proangiogenic factors in the developing brain29,30. On the contrary, neurons and astrocytes support neoangiogenesis at mature stages and adulthood through the secretion of vascular endothelial growth factor31,32. These changes in the brain cytoarchitecture, which are reflected in brain organoids too, might lead to differential changes in the integration and response to CAM engrafting, depending on the maturation stage of brain organoids. Therefore, we grafted brain organoids at differentiation day 13 in vitro (DIV), when neuroepithelial cells and radial glia expand; at DIV 37, at the onset of neurogenesis; at DIV 60, when neurogenesis is robust and neurons of different identities are terminally differentiated; and at DIV 120, when a plethora of mature neurons is populating the organoid, generating a well-developed neuronal network and astrogenesis is also widespread.
In all cases, the samples were collected after 5 days of in ovo (DIO) incubation. To avoid biases due to batch effect in brain organoids, all comparisons between grafted and non-grafted organoids were performed in the same batch in a pairwise manner. Organoids were derived from at least two independent experiments per time point and a total of six independent differentiations. Grafting of multiple maturation stages was performed in parallel in each grafting experiment. Upon grafting, organoids continued the maturation process adjacent to the CAM vessels without any evidence of infiltration of blood vessels into the organoid (Figure 2).
Organoid survival upon grafting ranges from 80% in the youngest and drops to ~66% in mature organoids after 5 days post grafting, suggesting that permissivity and organoid grafting plasticity might change over time (Table 1). The organoid viability was determined by the presence of pyknotic nuclei in H&E stainings. The number of pyknotic nuclei was similar between transplanted and the non-transplanted control organoids (Figure 3), suggesting that CAM is permissive to sustain the live organoid upon transplantation. Moreover, no histological or hemogenic release was detected at the CAM, and the embryos remain alive, suggesting that neither the grafting procedure nor the organoid disrupt the integrity of the CAM or significantly affect embryo development.
Cell composition in grafted brain organoids
Unguided brain organoids have a marked inter- and intraorganoid cellular variability. However, they are predominantly neural, generating waves of progenitors, neurons, and astrocytes at a sequential pace2,26. Next, we evaluated whether grafting alters the neural composition in the brain organoids. Younger organoids (13 DIV + 5 DIO: 18 days) show no differences from their control non-grafted organoids in the presence of neurons (TUBB3+). At later maturation stages (60 DIV +5 DIO: 65 days and 120 DIV + 5 DIO: 125 days), the proportion of neurons (TUBB3+) is dramatically decreased compared to that in non-grafted organoids (Figure 4). In non-grafted organoids matured for 60 DIV, neurons are widely spread all around the organoids.
Notably, the number and localization of glial fibrillary acidic protein-positive (GFAP+) astrocytes, evidenced an increase compared to those observed in the non-grafted controls. Moreover, these GFAP+ astrocytes acquired an unexpected localization in the outer side of the organoid. In 18 DIV, free organoids (non-grafted), few astrocytes can be seen and no structural distribution could be detected; on the contrary, they appear homogeneously spread throughout the organoid (Figure 3).
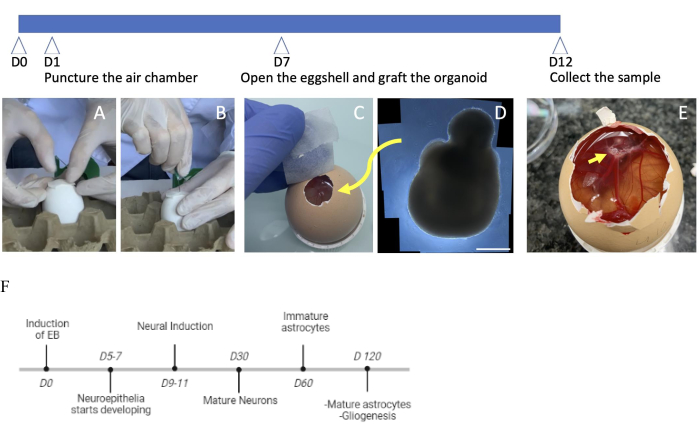
Figure 1: Timeline of CAM assay setup for organoid transplantation. (A) Surgical paper adhesive bandage application on top of the egg. (B) Making a hole in the shell with a needle over the adhesive bandage. (C) Open the eggshell's window, view from the top, ready for the transplantation of a (D) human brain organoid (day 60). Scale bar = 100 µm. (E) Visualizing the engrafted organoid at the CAM after 5 days of in ovo incubation. (F) Hallmarks of brain organoid development from Day 0 to Day 120. The arrow indicates the location of the organoid. Abbreviation: CAM = chorioallantoic membrane. Please click here to view a larger version of this figure.
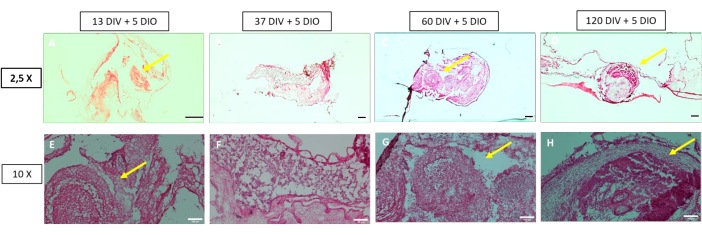
Figure 2: Hematoxylin and eosin staining of grafted organoids in the chorioallantoic membrane. (A) 13 DIV + 5 DIO organoid; (B) 37 DIV + 5 DIO organoid; (C) 60 DIV + 5 DIO organoid; and (D) 120 DIV + 5 DIO organoid; red squares mark the organoid. Scale bar = 500 µm. Abbreviations: DIV = day in vitro; DIO = day in ovo. Please click here to view a larger version of this figure.
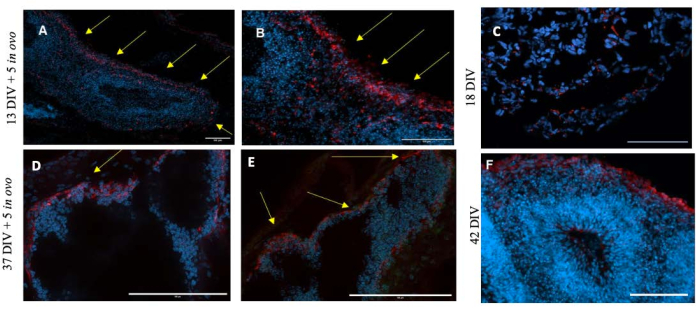
Figure 3: Immunocytochemistry of organoids at different stages of development. GFAP was used for staining and DAPI for the nuclei. Scale bar = 100 µm. (A,B) Images of the same 13 DIV + 5 DIO grafted organoid where astrocytes are seen around the edge of the organoid. (C) Image of 18 DIV free organoid where astrocytes are barely visible. The difference between free and grafted organoids is very noticeable. (D,E) Images of 37 DIV + 5 DIO grafted organoid where few astrocytes can be seen. (F) Image of 42 DIV free organoid. Yellow arrows indicate the location of the astrocytes. Abbreviations: DIV = day in vitro; DIO = day in ovo; DAPI = 4',6-diamidino-2-phenylindole; GFAP = glial fibrillary acidic protein. Please click here to view a larger version of this figure.
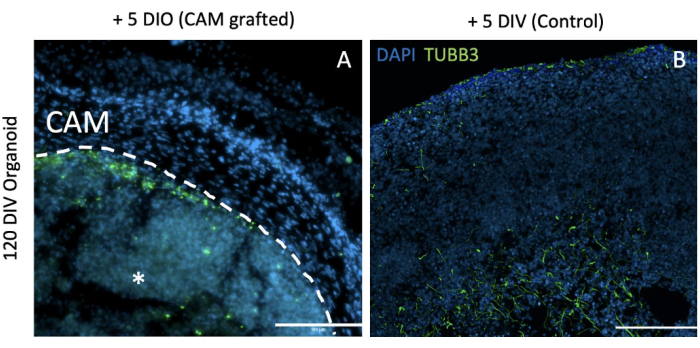
Figure 4: Immunocytochemistry of 120 DIV organoids. TUBB3 was used for staining the neurons and DAPI for the nuclei. Scale bar = 100 µm; images were taken at 20x. (A) A 120 DIV +5 DIO grafted organoid where few neurons can be seen. (B) A 125 DIV, free organoid with plenty of neurons. The asterisk indicates the location of the organoid while the dashed line represents the limit between the organoid and the CAM (determined by histological analysis). Abbreviations: DIV = day in vitro; DIO = day in ovo; DAPI = 4',6-diamidino-2-phenylindole; CAM = chorioallantoic membrane. Please click here to view a larger version of this figure.
| Organoids | 13 DIV | 37 DIV | 60 DIV | 120 DIV |
| Alive | 6 (75%) | 10 (83%) | 6 (66%) | 6 (66%) |
| Dead | 2 (25%) | 2 (16%) | 3 (34%) | 3 (34%) |
Table 1: Embryonic survival rates after CAM organoid grafting. Abbreviations: DIV = day in vitro; DIO = day in ovo; CAM = chorioallantoic membrane.
