For the representative results we present data from control C57BL/6J adult mice that received the neutral stimulus pairings with the unconditioned stimulus (shock condition) compared to mice that received the neutral stimulus but did not receive the unconditioned stimulus (no shock condition). It is important to run this condition when first setting up this behavioral test to determine whether the protocol has been performed correctly.
The data in Figure 1 represent the training day of C57BL/6J mice in the trace fear conditioning test. It is not generally necessary to compare the groups across all conditions during the training day. However it is useful to examine the baseline period to evaluate whether there are initial differences in baseline. We did not observe any differences in freezing levels in the baseline condition t(1,10) = 0.6, p = 0.56. This demonstrates that there are no initial differences in freezing levels. In general there are low freezing levels at the early learning trials but there is an increase in freezing behavior over the conditioning pairing trials. When we performed a mixed-model ANOVA that analyzed differences between the Shock and No Shock condition over the 16 periods we found a main effect of group F(1,10) = 60.3, p < 0.001. There was a main effect of group F(1,10) = 215.9, p < 0.001, and a group by time interaction F(1,10) = 133.9, p < 0.001. The analysis demonstrates that there was a significance difference in acquisition between the two groups.
The data for Figure 2 illustrate the freezing behavior between the mice that had tone shock pairings (Shock Condition) and the mice that received the same procedure without the shock pairing (No Shock Condition) across the 5 tone conditioning trials. The mice in the Shock Condition formed an association between the tone shock pairings in trace fear conditioning. A mixed-design ANOVA was used to examine the main effect of learning for group F(1, 10) = 83.48, p < 0.001. There was also a main effect of conditioning trials (time) F(3,30) = 24.83, p < 0.001, and group X time interaction F(3,30) = 4.7, p < 0.01. Since there was a group x time interaction, separate t-tests per time point were conducted to examine the difference between the groups at each time point. Separate t-tests reveal significant difference between the groups at the baseline t(1,10) = 6.8, p < 0.001; tone t(1,10) = 8.6, p < 0.001; trace period t(1,10) = 5.3, p < 0.001, and in the inter-trial interval t(1,10) = 5.1, p < 0.001. For the tone, trace period, and the inter-trial interval the data was averaged across the three presentations. The data demonstrate that the trace conditioning experiment successfully produced learning differences to the trace conditioning protocol.
The data for Figure 3 demonstrate the freezing behavior between the mice that had tone shock pairings (Shock Condition) and the mice that received the same procedure without the shock pairing (No Shock Condition) in the context condition. The context condition testing was presented 48 hr after trace conditioning is implemented. The mice in the Shock Condition had significantly more freezing in the original context than the No Shock Condition mice F(1,10) = 12.5, p < 0.01. There was also a main effect of time F(7,70) = 5.5, p < 0.001; however, there was no interaction between time and group F(7,70) = 0.78, p = 0.61. The data demonstrate that the trace conditioning experiment successfully produced contextual learning when the CS was paired with an US in trace fear conditioning.
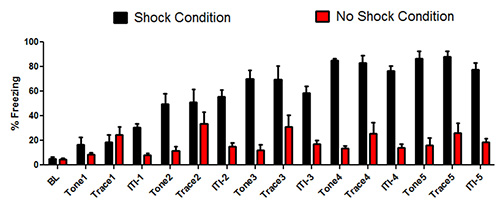
Figure 1. Data from the training day of trace fear conditioning. The black bars represent the data from the mice that received the aversive unconditioned stimulus (shock). The red bars represent mice that did not receive the aversive unconditioned stimulus (no shock) but received the tone stimuli. The bars represent the mean ± Standard error of the mean (SEM) percent freezing for mice during the training day.
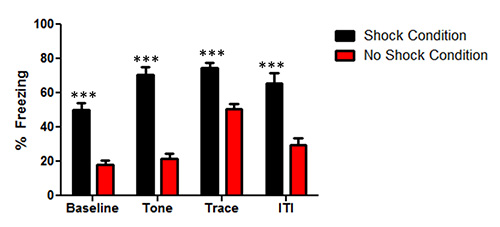
Figure 2. Data from mice in the tone test following trace fear conditioning. The mice that received the unconditioned stimulus (Shock condition) had more freezing compared to the mice that did not receive the shock (No Shock Condition) in the baseline, tone, trace, and intertrial interval (ITI). The bars represent the mean ± Standard error of the mean (SEM) percent freezing for mice during the tone test. Astericks (***) indicate a significant group difference (p < 0.001).
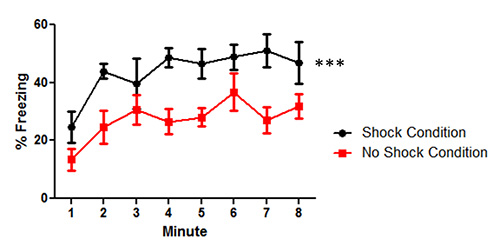
Figure 3. Data from mice tested in the context test following trace fear conditioning. The mice that received the shock had more freezing compared to the mice in the No Shock condition in the context test condition across the 8 min trial. The data points represent the mean ± Standard error of the mean (SEM) percent freezing for mice during the context test. Astericks (***) indicate a significant group difference (p < 0.001).
