Using non-denaturing PAGE, we analyzed the oligomeric state of the human wild type MxA, the dimeric interface mutants MxA(R640A) and MxA(L617D) as well as the monomeric interface mutant MxA(M527D) from cell lysates 12. Cells were lysed in a buffer containing 1% octylphenoxypolyethoxyethanol (NP-40) and iodoacetamide to ensure protein solubilization and protection of free thiol groups (see Figure 1). As described before, salt and small metabolites were removed by dialysis 19. Protein separation was carried out by non-denaturing PAGE. To facilitate efficient western blotting, the gel was incubated in SDS buffer before blotting. The MxA proteins were visualized by immunostaining using a rabbit polyclonal antibody directed against MxA. The workflow is described in Figure 2.
To compare the oligomeric state of endogenous human MxA protein from IFN-α stimulated A549 cells, we transfected Vero cells (lacking endogenous MxA) with recombinant wildtype, monomeric and dimeric MxA variants. These recombinant wild type, monomeric and dimeric MxA variants formed stable tetramers, monomers and dimers, respectively, when compared to an unstained native protein marker (Figure 3A). Therefore, we used these recombinant proteins to assess the oligomeric state of endogenous human MxA protein derived from IFN-α stimulated A549 cells. Figure 3B reveals that the size of MxA in lysates of IFN-α-stimulated A549 cells corresponds to a tetramer.
Taken together, we describe a method to determine the oligomeric state of the human MxA protein from cell lysate. Our non-denaturing PAGE approach can also be used to assess the oligomeric state of other oligomeric protein complexes.
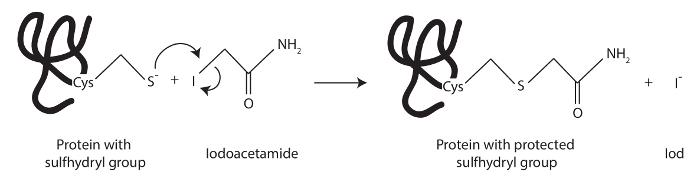
Figure 1: Structure and reaction scheme of iodoacetamide. Iodoacetamide irreversibly protects the thiol group of free cysteins by forming a thioether bond. This stable modification results from the nucleophilic substitution of the iodine with the sulfur atom from the cystein. Please click here to view a larger version of this figure.
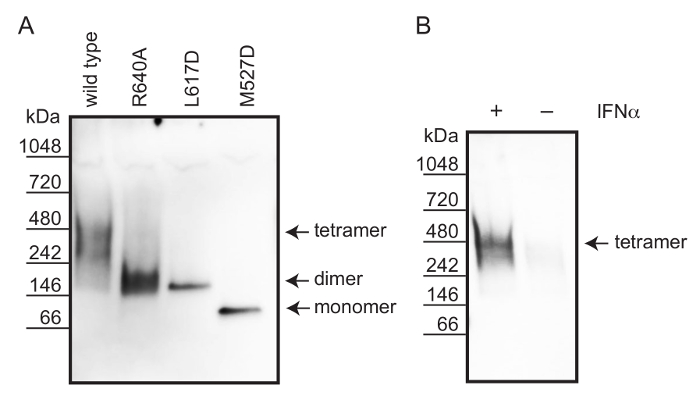
Figure 2: Workflow diagram of non-denaturing PAGE. A systematic representation of the non-denaturing PAGE approach of cell lysates. During cell lysis, detergents solubilize the proteins and thiol groups are protected by iodoacetamide to prevent protein aggregation. Dialysis removes small metabolites and salts that could interfere with non-denaturing PAGE 19. The complex separation is performed under non-denaturing conditions. Detection of the oligomeric complexes is achieved by Western blot followed by immunostaining. Please click here to view a larger version of this figure.
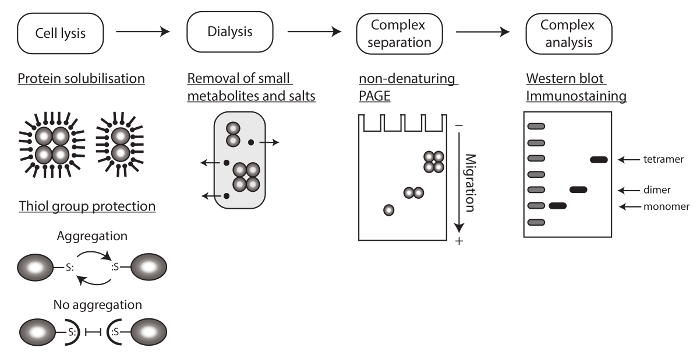
Figure 3: Determination of the oligomeric state of the human MxA protein using non-denaturing PAGE and Western blotting. (A) Recombinant MxA variants ectopically expressed in Vero cells. The complexes of wild type MxA (tetramer) interface mutants MxA(R640A), MxA(L617D) (dimers) and MxA(M527D) migrate at their expected molecular weights, confirming their oligomeric state. (B) A549 cells were stimulated with 1,000 IU per ml of IFN-α to induce MxA expression. The endogenous MxA shows a band that corresponds to the tetrameric form. Please click here to view a larger version of this figure.
| Buffer name | Content | Comments | ||
| Lysis buffer | 20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 5 mM MgCl2, 100 μM iodoacetamide, 50 mM NaF, 1 mM Na3VO4, 1% NP-40, 50 mM β-glycerophosphate, 1 tablet per 50 ml Lysis buffer of EDTA free, protease inhibitor cocktail |
Add iodacetamide, NaF, Na3VO4, octylphenoxypolyethoxyethanol (NP-40), β-glycerophosphate and protease inhibitor cocktail right before cell lysis Iodacetamide is light-sensitive and instable when in solution. Prevent light exposure and dissolve right before use. |
||
| Dialysis buffer | 20 mM Tris-HCl (pH 6.8), 10% glycerol, 0.1% CHAPS, 0.5 mM DTT |
CHAPS can be raplaced by other non-denaturing detergents | ||
| Running buffer | 25 mM Tris-HCl (pH 8.3), 192 mM glycine, 0.1% CHAPS, 0.5 mM DTT |
At pH 8.3, most proteins are negatively charged. However, for basic proteins, an acidic pH should be used. Otherwise, the proteins will run into the opposite direction and will be lost. CHAPS can be replaced by other non-denaturing detergents |
||
| Sample buffer | 310 mM Tris-HCl (pH6.8), 0.05% bromophenol blue, 50% glycerol |
|||
| SDS buffer | 25 mM Tris-HCl (pH 8.3), 192 mM glycine, 0.1% SDS |
|||
| Blotting buffer | 25 mM Tris, 192 mM glycine, 20 % Methanol |
For very large complexes, Methanol can be ommitted | ||
| Blocking buffer | 50 mM Tris-HCl (pH 7.4) 150 mM NaCl 0.05 % Tween 20 5 % Milk powder |
|||
| Growth medium | Dulbecco's modified medium 1x Penicillin/Streptomycin 2 mM Glutamine 10% Fetal Calf Serum |
|||
Table 1: Buffer recipes required for non-denaturing PAGE.
