Using the described method results in serial sections stained by the eye pigment33 that encompass the entire fly head. A part of this is shown in Figure 1B, where the sections from an individual head are shown from top to bottom. The sections from different flies are seen left to right in this example. To facilitate orientation and identification of the flies, an eyeless fly (sine oculis) is inserted as a marker at position 3 (arrow, Figure 1B).
To quantify neurodegeneration, we measure the formation of vacuoles that can be detected in these sections. Vacuoles are defined as round, dark spots that are within the green fluorescent neuropil (arrowheads in Figure 2 and 3) or cortex and that are visible in at least 2 consecutive sections of the fly brain. Quantifying neurodegeneration by measuring vacuoles can either be done by focusing on a specific brain region or by analyzing the entire brain. Limiting the analysis to a specific region of the brain is useful in cases where a mutation only affects a specific region, like the olfactory lobes in the futsch'olk mutant34, but it can also be used when there is severe degeneration in all or many regions of the brain. An example for the latter is the swiss cheese (sws) mutant (Figure 2), where measuring all vacuoles would be too time consuming. We therefore took only one image and, to ensure that the measurements were always done at the same level, we took all images at the level of the great commissure (gc, Figure 2A), which is only contained in one or two sections. Whereas we did not detect vacuole formation in 1-day-old sws'1 flies (data not shown), a loss-of-function allele35, some vacuoles were detectable in 7-day-old sws'1 flies (arrowheads, Figure 2A). Aging the flies to 14 d (Figure 2B) and 21 d (Figure 2C) further increased this phenotype, showing its progressive nature. Counting the number of vacuoles in the deutocerebral neuropil (dn) using the described method confirmed a significant increase in the number of vacuoles with age. Also, the combined area encompassed by vacuoles was significantly increased with aging (Figure 2D).
However, not all mutants show such a severe phenotype as sws, and in those cases, differences in degeneration are difficult to determine when focusing on a small area. Similarly, the degeneration that occurs during aging is quite mild (Figure 3A – C) and therefore, we analyzed the entire brain when quantifying this phenotype. Determining the sum of all vacuoles in the brain revealed a significant increase with age, and this was also the case when measuring the combined area of these vacuoles (Figure 3D).
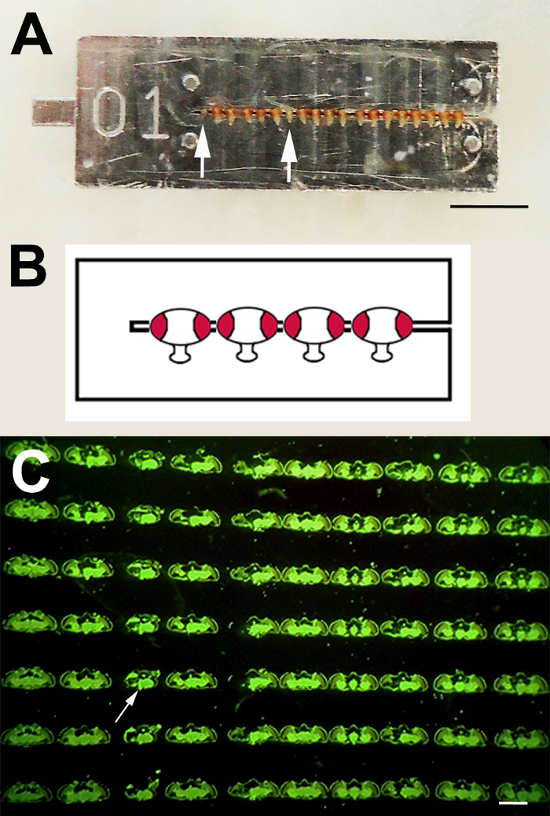
Figure 1. Paraffin Serial Sections. A) Using the collar method, experimental and control flies can be processed as one sample by threading them onto one collar. Eyeless sine oculis flies are inserted for orientation (arrows). B) Schematic showing the orientation of the fly heads in the collar. C) In this image, sections from different fly heads are oriented left to right on the slide. From top to bottom on the slide, serial sections from the same fly head can be seen. In this case, a sine oculis fly was inserted at position three (arrow). The sections are stained by the fluorescent eye pigment that washes over the sections after cutting. Scale bar in A = 5 mm and in C = 0.5 mm. Please click here to view a larger version of this figure.
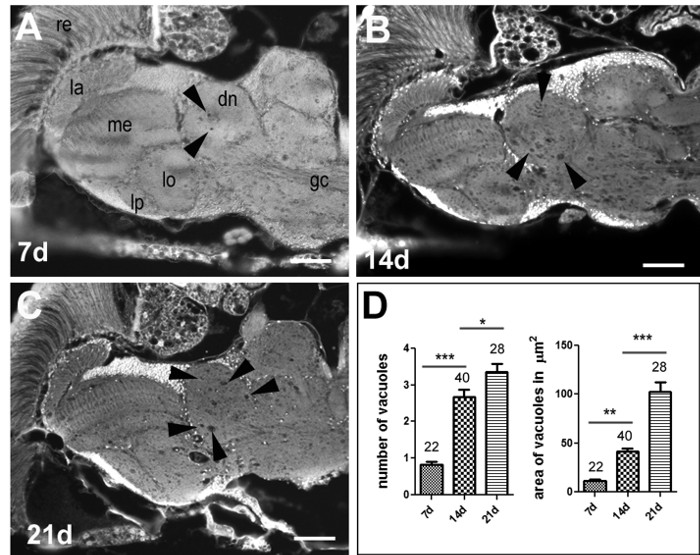
Figure 2. Progressive Neurodegeneration in the Swiss-cheese Mutant. Paraffin head section from 7-day- (A), 14-day- (B) and 21-day- (C) old sws'1 flies. The arrowheads point to vacuoles that have developed with aging. The age-related degeneration is quantified by counting the number of vacuoles and measuring their combined area (D). The SEM and number of analyzed flies is indicated. The scale bar = 25 µm. ***p <0.001. Please click here to view a larger version of this figure.
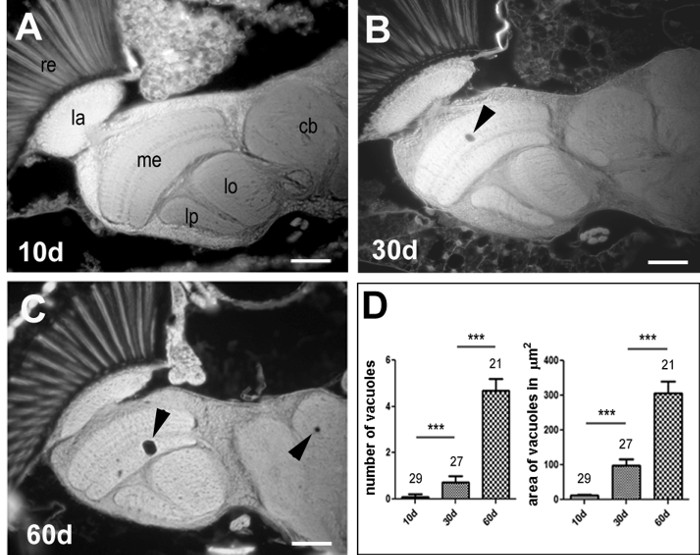
Figure 3. Neurodegeneration Occurs with age. Paraffin head section from 10-day- (A), 30-day- (B) and 60-day- (C) old wild-type flies. The arrowheads point to vacuoles that have developed in aged flies. The age-related degeneration is quantified by counting the number of vacuoles and measuring their combined area (D). The SEM and number of analyzed flies is indicated. The scale bar = 25 µm. ***p <0.001. Please click here to view a larger version of this figure.
