For proper maintenance and use of any bacterial strain, it is recommended to know the doubling time of the strain(s) of interest. Herein, the varying growth rates of commonly used V. cholerae strains were demonstrated through a growth curve, and approximate doubling times were calculated using linear regression. WT El Tor N16961 and El Tor variant MQ1795 demonstrated shorter doubling times (~1 h and ~1 h, respectively) than WT classical O395 (~2 h) (Figure 1; Table 2).
V. cholerae genetic manipulation and subsequent analysis often relies on the ability to properly distinguish between biotypes. PCR based genetic screens and phenotypic assays were collectively implemented as a reliable system for distinguishing between biotype backgrounds of V. cholerae clinical and environmental isolates; for representation, biotype reference strains (WT classical O395, WT El Tor C6706, and WT El Tor N16961) and representative El Tor variants (MQ1795 and BAA-2163) were included (Table 1). WT classical O395 demonstrated classical ctxB and tcpA sequences. Conversely, WT El Tor strains N16961 and C6706 demonstrated El Tor ctxB and tcpA sequences. Interestingly, MQ1795 and BAA-2163 contained the classical biotype ctxB subunit comparable to O395, yet both El Tor variants contained the tcpA indicative of the El Tor biotype background (Table 1). WT classical biotype strain O395 showed sensitivity to polymyxin B, while WT El Tor biotype strains (C6706 and N16961) showed resistance and exhibited growth on LB agar plates supplemented with polymyxin B. The representative El Tor variant strains (MQ1795 and BAA-2163) demonstrated similar resistance to the antibiotic relative to the WT El Tor biotype strains (C6706 and N16961) (Figure 2; Table 2). WT classical biotype strain O395 did not grow on minimal citrate media, while WT El Tor strains (C6706 and N16961) were able to utilize citrate as a carbon source and exhibit growth on minimal citrate media. Representative El Tor variant biotype strains (MQ1795 and BAA-2163) demonstrated growth comparable to that of the WT El Tor biotype strains (C6706 and N16961) (Figure 3; Table 2). WT classical strain O395 and WT El Tor strain N16961 possess a non-functional HapR, and, thus did not demonstrate HapR-regulated protease activity; WT El Tor strain C6706, and representative El Tor variants (MQ1795 and BAA-2163) are hapR-positive-visualized as a zone of clearance emanating from the point of inoculation (Figure 4; Table 2). WT classical strain O395 does not secrete hemolytic enzymes and was therefore γ-hemolytic, while WT El Tor biotype strains (C6706 and N16961) and representative El Tor variant strains (MQ1795 and BAA-2163) secrete hemolytic enzymes that completely lyse red blood cells surrounding the point of inoculation and showed β-hemolysis (Figure 5; Table 2). Motility varies across, and within, biotype strains; however, WT El Tor strain N16961 and El Tor variants (MQ1795 and BAA-2163) demonstrated hyper-motility when compared to the relatively less motile WT classical strain O395 and WT El Tor strain C6706 (Figure 6; Table 2). WT classical strain O395 and representative El Tor variants did not metabolize glucose to produce acetoin, while WT El Tor biotype strains produced acetoin as a byproduct of glucose fermentation, as indicated by development of a deep red color during the Voges-Proskauer assay (Figure 7; Table 2).
| Strain | ctxB Gene | tcpA | Reference | ||
| Base 115 | Base 203 | Biotype | Biotype | ||
| O395 | C | C | classical | classical | 8 |
| C6706 | T | T | El Tor | El Tor | 8 |
| N16961 | T | T | El Tor | El Tor | 8 |
| MQ1795 | C | C | classical | El Tor | 13 |
| BAA-2163 | C | C | classical | El Tor | 8 |
Table 1: Biotype Dependent Genetic Distinctions of Vibrio cholerae Reference Strains. Shown in this table are the DNA base changes and relative positions in the genes ctxB and tcpA. WT classical O395 and WT El Tor strains N16961 and C6706 are commonly used biotype reference strains. MQ1795 and BAA-2163 are known El Tor variants.
| Assay | Application | Medium Selection | Incubation | Expected Results | ||||
| O395 | C6706 | N16961 | MQ1795 | BAA-2163 | ||||
| 2.3) Growth Curve27 | Determines doubling times of various V. cholerae strains | 1.2) Liquid LB Broth | up to 30 h (37 °C with aeration) | ~2 h | ND* | ~1 h | ~1 h | ND* |
| 3.1) PCR Based Genetic Screen using ctxB and tcpA8 | Differentiates between classical and El Tor biotype backgrounds of ctxB | N/A | N/A | classical | El Tor | El Tor | classical | classical |
| Differentiates between classical and El Tor biotype backgrounds of tcpA | N/A | N/A | classical | El Tor | El Tor | El Tor | El Tor | |
| 3.2) Polymyxin B Resistance21 | Sensitivity to the antibiotic polymyxin B | 1.5) LB agar plates supplemented with polymyxin B | 18 h (37 °C) | – | + | + | + | + |
| 3.2) Citrate Metabolism22 | Ability to metabolize citrate as sole carbon source | 1.6) Minimal citrate medium agar plates | 24 h (37 °C) | – | + | + | + | + |
| 3.2) Casein Hydrolysis23 | HapR-regulated protease activity | 1.7) Milk agar plates | 18 h (37 °C) | – | – | + | + | + |
| 3.2) Hemolysis23 | Measures hemolytic activity | Blood agar plates | 48 h (37 °C) | Gamma | Beta | Beta | Beta | Beta |
| 3.3) Motility23 | Measures degree of motility | 1.8) Motility agar plates | 14-24 h (37 °C) | 10 mm | 15 mm | 21 mm | 25 mm | 29 mm |
| 3.4) Voges-Proskauer21 | Measures ability to ferment glocose and produce acetoin as a byproduct | 1.9) Liquid Voges-Proskauer medium | up to 4 h (room temperature) | – | + | + | – | – |
| Note: "ND*" denotes not determined; "+" denotes a positive result; "-" denotes a negative result | ||||||||
Table 2: Summary of Genetic and Phenotypic Assays Used for Vibrio cholerae Biotype Distinction. This table summarizes the various genetic and phenotypic assays, applications, and expected results collectively used to differentiate between classical and El Tor biotypes used in this study. Protocol numbers and references to specific protocols are indicated in the Assay column. "ND*" denotes Not Determined; "+" denotes a positive result; "-" denotes a negative result.
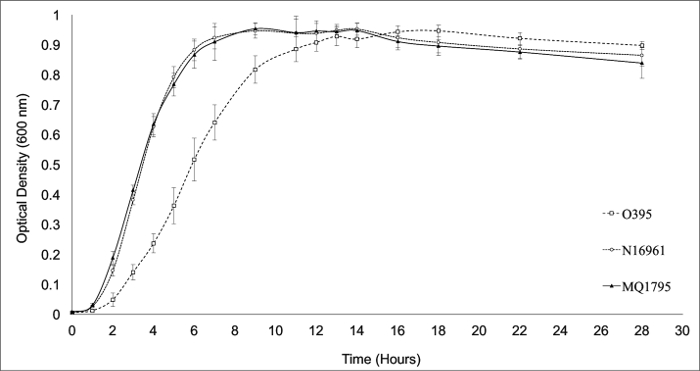
Figure 1: V. cholerae Growth Curve of WT Classical O395, WT El Tor N16961, and El Tor Variant MQ1795. Growth rates of biotype reference strains (WT classical O395 and WT El Tor N16961) and representative El Tor Variant MQ1795, grown in LB broth with aeration at 37 °C, were analyzed by measuring the OD600 every hour beginning at T0. Growth curves were performed on 8 independent experimental replicates, with each replicate representing an independent culturing event for each trial. WT El Tor strain N16961 and El Tor variant MQ1795 demonstrated shorter doubling times (~1 h and ~1 h, respectively) relative to WT classical strain O395 (~2 h), as observed by a longer doubling time. Please click here to view a larger version of this figure.
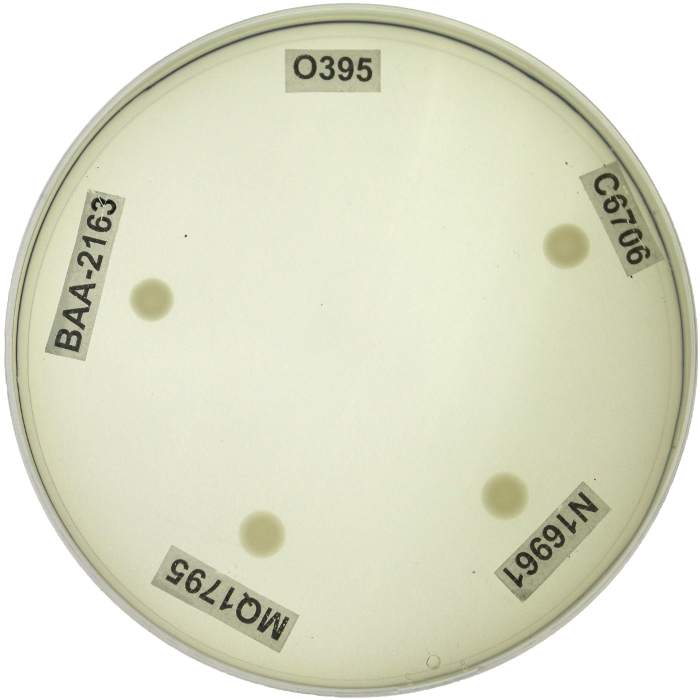
Figure 2: Determining Polymyxin B Resistance Using LB Agar Supplemented with Polymyxin B. Resistance to the peptide antibiotic polymyxin B was determined by the ability to grow on LB agar supplemented with 50 IU/µL polymyxin B. WT classical strain O395 showed no growth on agar supplemented with polymyxin B and was considered sensitive to the antibiotic. While WT El Tor strains (C6706 and N16961) and representative El Tor variants (MQ1795 and BAA-2163) exhibited growth in the presence of the antibiotic and were considered resistant to polymyxin B. Plates were incubated at 37 °C for 18 h. Please click here to view a larger version of this figure.

Figure 3: Measuring Citrate Metabolism Using Minimal Citrate Media. The ability to utilize citrate as a sole carbon source was determined by the isolate's ability to grow on minimal citrate media. WT classical strain O395 did not grow on minimal citrate media (negative). Growth was evident by all WT El Tor strains (C6706 and N16961) and representative El Tor variants (MQ1795 and BAA-2163), which demonstrated the ability to utilize citrate as a sole carbon source (positive). Plates were incubated at 37 °C for 18 h. Please click here to view a larger version of this figure.
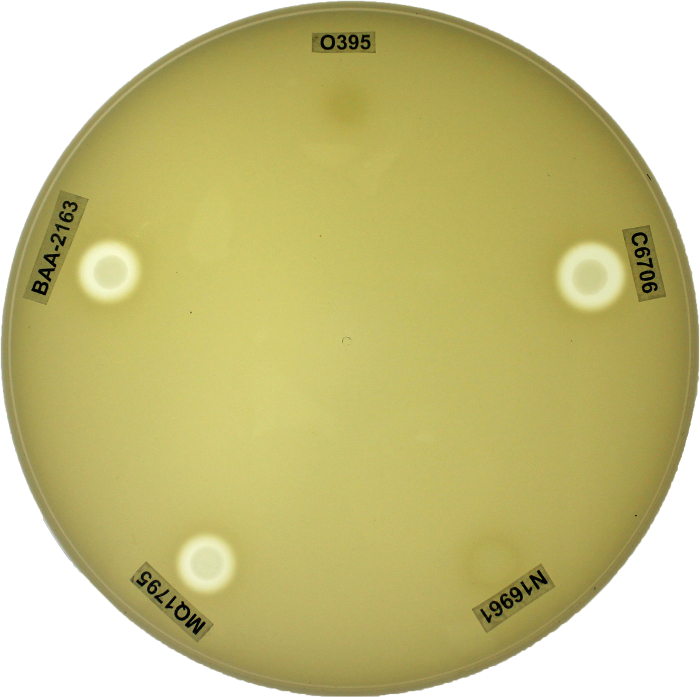
Figure 4: Measuring HapR-regulated Proteolytic Casein Hydrolysis Using Milk Agar Media. Casein hydrolysis through HapR-regulated protease activity was determined by a visual zone of clearance surrounding the point of inoculation on milk agar. Strains containing a non-functional HapR, such as WT classical strain O395 and WT El Tor strain N16961, did not produce a zone of clearance surrounding the point of inoculation (hapR-negative). WT El Tor strain C6706 and representative El Tor variants (MQ1795 and BAA-2163) contain a functional HapR, which can be visualized as varying sized zones of clearance (hapR-positive). Plates were incubated at 37 °C for 18 h. Please click here to view a larger version of this figure.

Figure 5: Measuring Hemolytic Activity Using Blood Agar Media. Hemolytic activity was measured using agar plates supplemented with sheep's blood. WT classical strain O395 does not secrete enzymes that lyse red blood cells (γ-hemolytic). WT El Tor strains (N16961 and C6706) and representative El Tor variants (MQ1795 and BAA-2163) secrete hemolytic enzymes, which resulted in a translucent zone of clearance surrounding the point of inoculation (β-hemolytic). Plates were incubated at 37 °C for 48 h. Please click here to view a larger version of this figure.
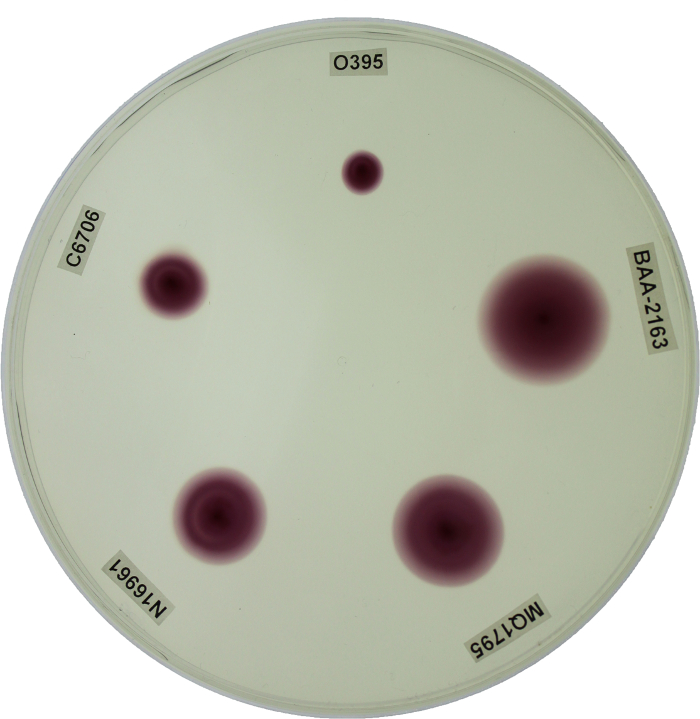
Figure 6: Determining Motility Using Motility Agar Plates. The zone of motility was indicated by a visual color change of the salt TTC which turns from clear to red when metabolized, indicating where the bacteria have moved. WT classical strain O395 (10 mm) and WT El Tor strain C6706 (15 mm) demonstrated minimal motility, while El Tor strain N16961 (21 mm) and representative El Tor variants (MQ1795 (25 mm) and BAA-2163 (29 mm)) demonstrated hyper-motility relative to O395 and C6706. Plates were incubated at 37 °C for 18 h. Please click here to view a larger version of this figure.
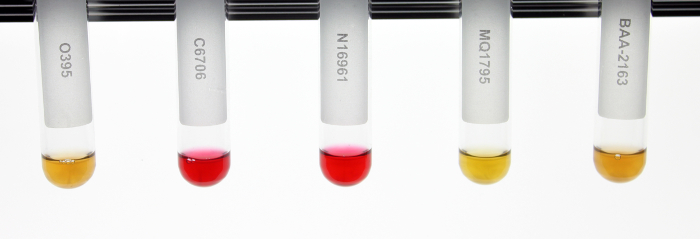
Figure 7: Voges-Proskauer Assay. Acetoin production via glucose fermentation was determined using the Voges-Proskauer assay. WT classical strain O395 and representative El Tor variants (MQ1795 and BAA-2163) did not produce acetoin as a result of glucose fermentation (negative). WT El Tor strains (N16961 and C6706) produced the byproduct acetoin and can be visualized by a deep red color change (positive). Tubes were incubated at room temperature for 4 h. Please click here to view a larger version of this figure.
