Bone culture can be performed starting from different stages. In Figure 1A-D, a comparison between cultured tibia and freshly extracted ones at equivalent stages is shown. The first observation is that up to two days of culture the size achieved is comparable to the in vivo bone growth for both cartilage and mineralized bone (Figure 1A,B,D). Longer culture periods lead to bigger differences between the cultured bones and the freshly extracted (Figure 1C). Additionally, as mentioned in the protocol, it is crucial to remove the soft tissue connecting both ends of the bones, as otherwise the bones will bend. Figure 1E shows an example of a tibia grown with incomplete removal of soft tissue versus a tibia without soft tissue.
Next, tibias were cultured for 2 days and their length was measured. As can be seen in Figure 2A,B, the measurement of the total length and of the mineralized part can be performed with an image analysis software. As shown by De Luca et al.26, treatment with RA has a strong effect on the growth of the tibias already after 2 days of treatment and a similar result was observed in our cultures (Figure 2B-D). Importantly, the experiment was performed using paired bones, with the right bone as control and the left treated with RA (Figure 2A,B,E), which helps overcome the natural variability in bone size between different specimens. Thus, the culturing method described is suitable for assessing the effect of different compounds on bone growth.
Next, the growth rate of the bones after culture was assessed. Bones were extracted, measured and cultured at E15.5, 16.5 or P1, and fixed and measured again two days later. Both the total increase in length and the length of the mineralized part were measured (Figure 3C). An example of E15.5 tibia and femur before culture (Figure 3A) and at the end point of the experiment (Figure 3B) are shown. As can be observed from the graph and the table (Figure 3C,D), there is a consistent increase in the total length of the tibia, corresponding to an approximate increase of 9−29% from the initial length. This is less increase than the one observed in vivo; the main difference is likely due to the level of the proximal cartilage, bigger than the distal and less accessible to nutrients. Indeed, EdU labeling showed fewer positive cells in this region after culture compared to freshly extracted bones of equivalent stage (Figure 4A,C). The EdU incorporation in the distal tibia was similar in the cultured and freshly extracted bones (Figure 4B,D) The distal cartilage contributes approximately one third to the total growth of the tibia in vivo at this stage27, comparable to the growth rate observed in culture, so we propose that the analysis should be focused on this part of the bone. Additionally, we assessed the mineralization of the cultured bones, and observe almost no increase in length of this region. The difference might be due to the absence of vessel and osteoblasts invasion in the ex vivo culture. This suggests that studies of cartilage growth can extend for several days, while if the region of interest is the ossified part of the bone, the period of culture should be shorter. This observation was confirmed by keeping tibia in culture for a longer period (up to 6 days). As can be seen in Figure 5, the total length of the skeletal element increases substantially, while almost no increase in the mineralized region is observed (Figure 5C).
Overall, these results suggest that the culture of long bones can be used to analyze the effect of different factors on overall bone growth and particularly to assess cartilage dynamics. While well-established metatarsal cultures can also be used with these purposes, we submit that both types of cultures complement each other, given the intrinsic differences between metatarsals and the rest of long bones.
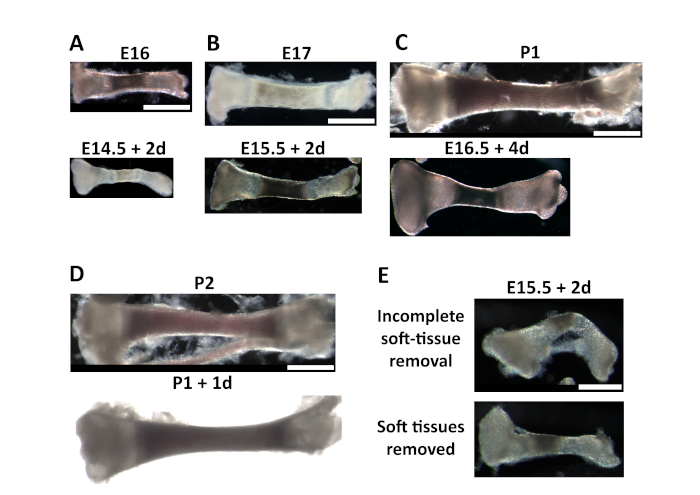
Figure 1: Example of cultured tibia for different time periods. (A-D) Comparison between freshly extracted tibia (top) and tibia extracted at E14.5 (bottom) and cultured for 2 days (A), extracted at E15.5 and cultured for 2 days (B), extracted at E16.5 and cultured for 4 days (C) and freshly extracted at postnatal day 2 (P2) and extracted at postnatal day 1 (P1) and cultured for 1 day (D). Note that, while cartilage growth remains quite physiological after different culturing periods, the ossified part shows a growth delay after culture time longer than 2 days compared to the freshly extracted bone at the corresponding stage. Scale bar = 1 mm. (E) Tibia cultured for 2 days starting at E15.5; note that the incomplete removal of the soft tissue between the two ends of the bones leads to the bending of the bone. Scale bar = 600 µm. Please click here to view a larger version of this figure.
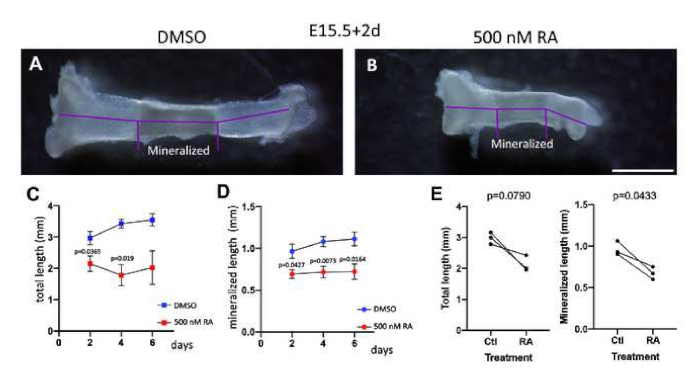
Figure 2: Measurement of the length of the bones upon retinoic acid (RA) treatment. (A-B) Tibias extracted at E15.5 and grown for 2 days with 0.1% DMSO (A) and RA (B). Note the difference in both total length and of the mineralized part. Scale bar = 1 mm. (C-D) Changes in total length (C) or the mineralized part (D) of the tibia over a period of 6 days in control situation and in presence of RA. Results are shown as mean ± standard deviation (SD); comparison is done by two-way analysis of variance (ANOVA) with the type of treatment as variable (p values are shown in the graph). (E) Comparison of the length of paired bones cultured for 2 days with either DMSO (right tibia) or RA (left tibia); each dot represents one of the 3 biological replicates per condition. Paired two-tailed Student’s t-test was used. Please click here to view a larger version of this figure.
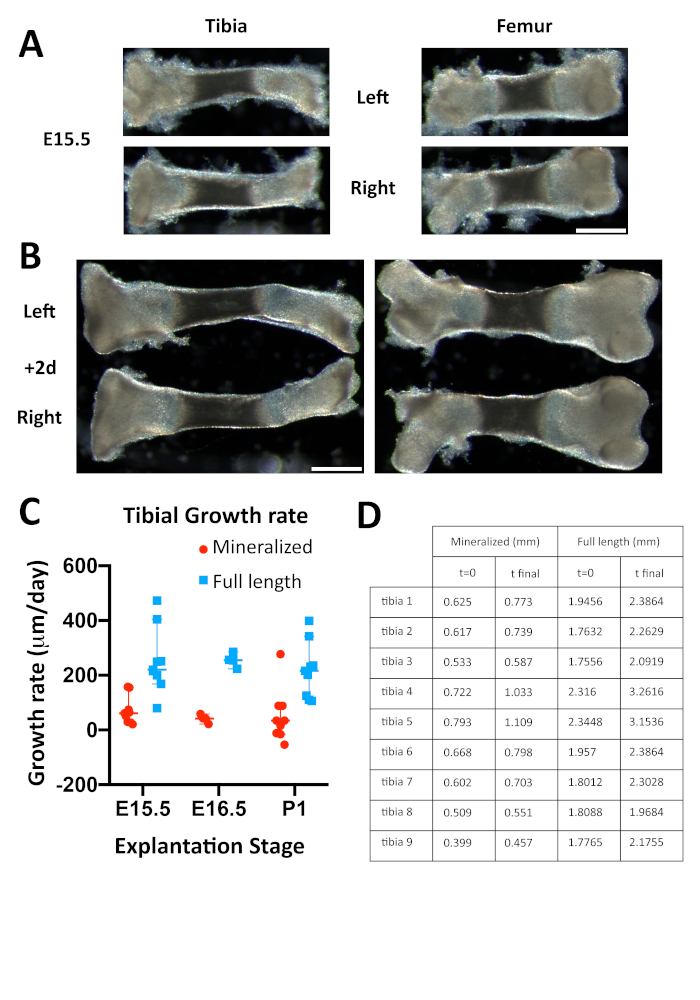
Figure 3: Comparison of total growth and mineralization of the tibia after 48 hours of ex vivo culture. (A) E15.5 tibias and femurs from left (top) and right (bottom) limbs prior to culturing. (B) The same tibias and femurs followed up after 2-day culture. Scale bar = 600 µm. (C) Graph representing the growth rate of tibias cultured at different developmental stages. Both the total growth and the growth of the mineralized part were assessed. (D) Table showing the initial and final length of the whole bone and the mineralized part before culturing at E15.5 and after 2 days in culture. Please click here to view a larger version of this figure.
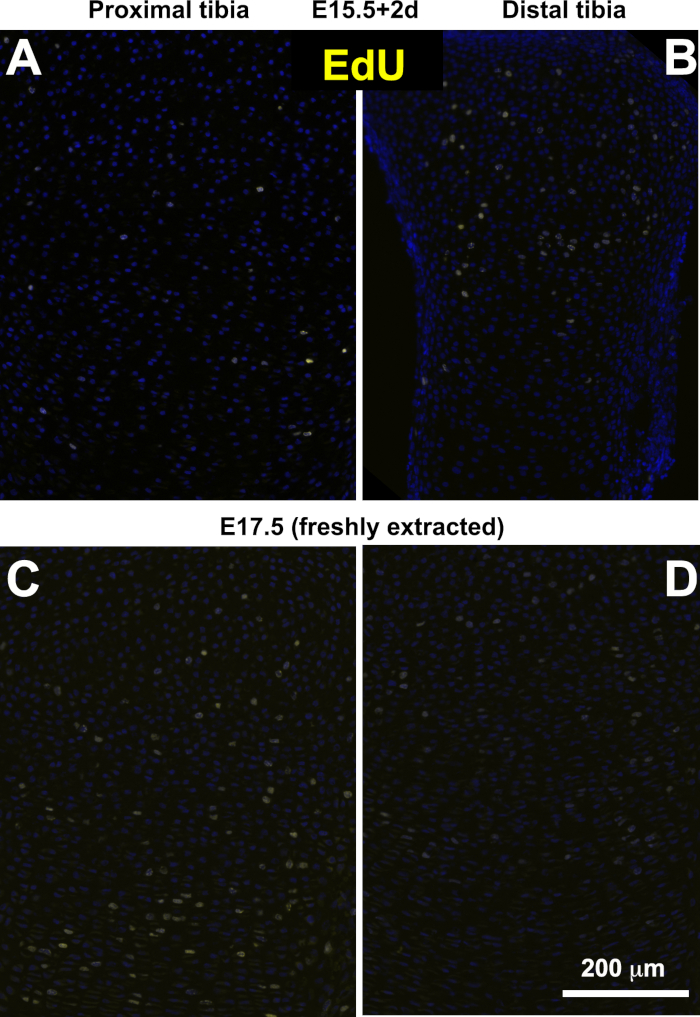
Figure 4: Bulky growth plates do not show much proliferation in culture. (A-B) EdU staining for proximal (A) and distal (B) tibial growth plates cultured from E15.5 for 2 days. (C-D) EdU staining for proximal (C) and distal (D) tibial growth plates freshly extracted at E17.5. Note the difference in the number of EdU(+) cells in the proximal tibia. Data include 6 cultured pairs of limbs and 3 freshly extracted. Please click here to view a larger version of this figure.
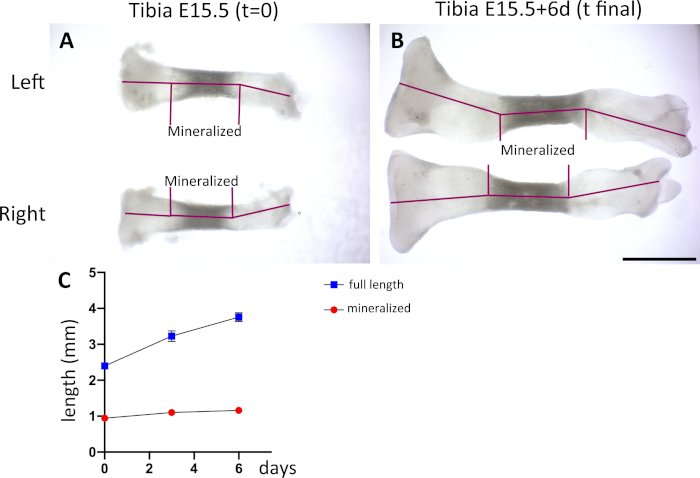
Figure 5: Longer periods of tibia culture show substantial cartilage growth but little mineralization. Freshly extracted tibia at E15.5 (t = 0) (A) and after 6 days in culture (B). Note the differences between cartilage growth and the growth of the mineralized part. Scale bar = 1 mm (C) Graph showing the changes in the total length and the mineralized region over a period of 6 days. n = 5 cultured tibias; SD at each time point is as follows: t = 0, full length = 0.0777, mineralized = 0.0213; t = 3 d, full length = 0.1495, mineralized = 0.056; t = 6 d, full length = 0.1193, mineralized = 0.0521. Please click here to view a larger version of this figure.
