In order to ensure that the appropriate imaging and analysis conditions are in place for the in situ evaluation of mitochondrial morphology in tissue, an in vitro exploration of mitochondrial morphology in response to a known modulator of mitochondrial morphology is recommended (Section 1). As an example, SNCA was genetically overexpressed in SH-SY5Y cells to induce changes in mitochondrial morphology as previously described26. Other insults that could be used as a control to worsen mitochondrial morphology would be starvation or the use of mitochondrial activity inhibitors such as MPP+. Cells were transfected and stained for α-synuclein (AS) to separate SNCA+ (AS+) and SNCA– (AS-) cells. They were also stained with TOMM2027 to visualize the mitochondrial network of cells. To make this analysis as similar as possible to that of a 5 µm tissue section, one confocal plane was analyzed as opposed to a maximum projection of multiple planes. Morphological analysis of one confocal plane of TOMM20 revealed that both the total number of mitochondria and their aspect ratio or AR (which correlates with the elongation of the organelle) were reduced in response to SNCA overexpression (Figure 1).
Immunostaining was performed for the mitochondrial protein VDAC1 in 5 µm paraffin-embedded mouse brain sections from animals injected with PFFs as described in the protocol section above. Substantia nigra pars compacta (SNc) dopaminergic neurons, which undergo degeneration in PD, were revealed through co-immunostaining with anti-tyrosine hydroxylase (TH) and were regionally separated from the ventral tegmental area and the substantia nigra pars lateralis. On the other hand, anti-phosphoSer129-α-synuclein (pS129) staining allowed us to discriminate cells that harbored pS129 lesions from healthy cells (pS129+ versus pS129-). SNc images of three different animals were taken, and subsequent image analysis of VDAC1 staining of TH-positive neurons revealed a reduction of both mitochondrial number counts and aspect ratio between neurons bearing pS129 lesions and neurons lacking these (Figure 2). These results indicate that the mitochondrial morphology of neurons harboring pS129 lesions is impaired compared to that of cells lacking pS129 lesions.
While this particular experiment shows a reduction in the AR, thereby highlighting a reduction in the elongation of mitochondria together with a reduction in global counts, which indicates a worsening of the mitochondrial morphology, the interpretation of the data should be experiment dependent. For example, a reduction in AR and counts can point to a global reduction in mitochondrial content as well as fragmentation, while a reduction in AR but an increase in global counts would point to a mitochondrial fragmentation phenotype. Therefore, it is important to interpret the data in the context of both measures.
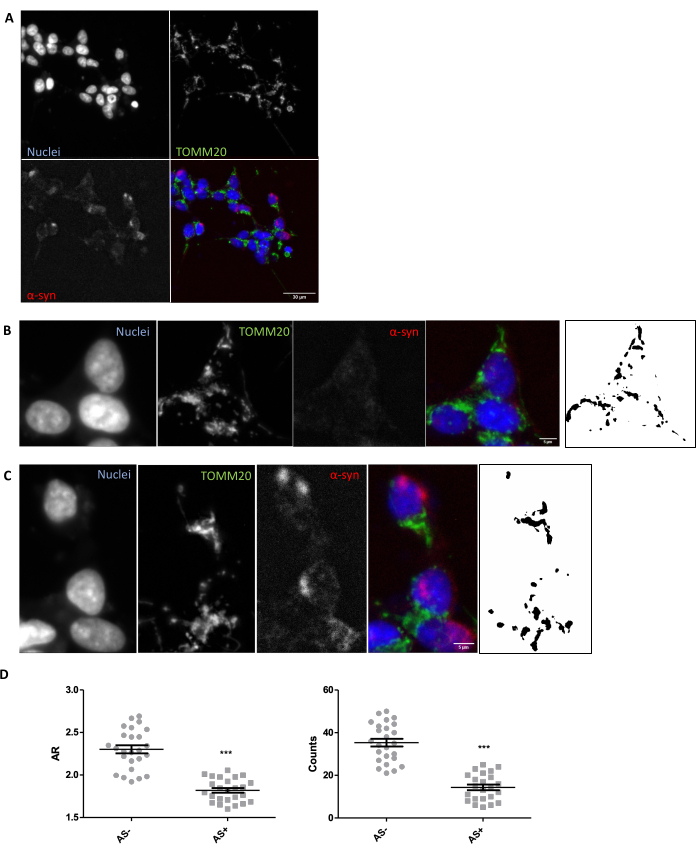
Figure 1: Mitochondrial morphology in an SNCA-overexpressing in vitro model. Co-immunostaining for TOMM20 (green), α-synuclein (AS, red), and DAPI (blue) on SNCA-overexpressing and not overexpressing (AS+ and AS-, respectively) cells (A). Detail of an AS- cell (B) and an AS+ cell (C). Black and white images in panels (B) and (C) represent the masks of the TOMM20 signal after applying the Fiji function described in the Protocol section. This mask enables quantification of the shape of the resulting structures. Mitochondrial counts and Aspect Ratio (AR) values of AS- and AS+ cells (N = 25 cells per condition) were quantified and represented as individual values as well as average ± SEM; **p-value < 0.05 t-test (D). Normality was assessed through the D'Agostino and Pearson normality tests. Scale bars: A, 30 µm; B,C, 5 µm. Please click here to view a larger version of this figure.
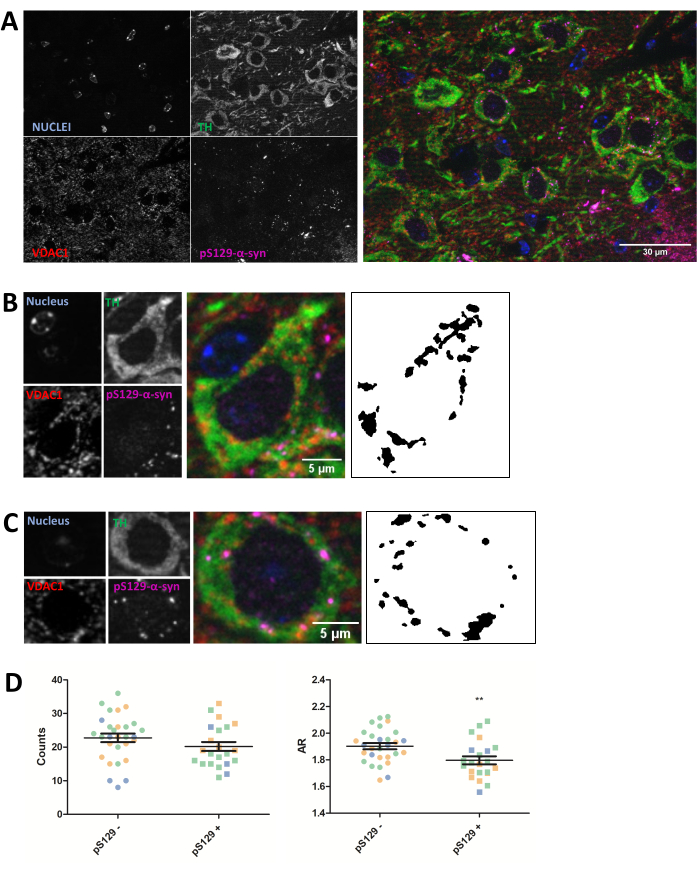
Figure 2: Mitochondrial morphology is affected in neurons harboring pS129 lesions. Co-immunostaining for TH (green), VDAC1 (red), phosphoS129-α-synuclein (magenta), and DAPI (blue) of the SNc of PFF-injected mice (A). The detail of a phosphoS129-α-synuclein-negative (pS129-) dopaminergic neuron (B) and of a phosphoS129-α-synuclein-positive (pS129+) dopaminergic neuron (C). Black and white images in panels (B) and (C) represent the masks of the TOMM20 signal after applying the Fiji function described in the Protocol section. This mask enables quantification of the shape of the resulting structures. Negative and positive cells were counted in samples from three different animals as illustrated by the different colors of the individual graphed values (blue, green, and orange). Mitochondrial counts and AR quantification of pS129- (N = 29) versus pS129+ (N = 22) in dopaminergic neurons were represented as average ± SEM as well as individual cell values; **p-value < 0.05 t-test (D). Normality was assessed through the D'Agostino and Pearson normality tests. Scale bars: A, 30 µm; B,C, 5 µm. Please click here to view a larger version of this figure.
