1. Microinjection procedure to label the zebrafish pronephros proximal tubule epithelium
For this procedure, we use a microinjection system (Harvard Appartus, PLI90), with pressurized air supplied by an air compressor (Jun-Air, Model 6-4), and micromanipulator apparatus (Narishege parts MN151, IP3, and GJI) that are assembled at a stereomicroscope station (Nikon, SMZ-Zoom 645) as per manufacturer instructions. Dextran conjugates are readily endocytosed by the nephron proximal tubule, enabling preferential labeling of this epithelial cell population9.
- Exposure to methylene blue can accentuate the autofluorescence of the yolk sac and makes visualization of the nephron tubules more difficult.
- Raise the fertilized embryos at 28.5°C to a developmental timepoint between 48-55 hours post fertilization (hpf). This is the ideal stage to perform larvae microinjections because of the ease at which intramuscular microinjection can be performed, and because renal function commences at this time.
- Remove the chorion from any unhatched zebrafish using a pair of fine forceps. Rinse the chorion debris from the petri dish and refill the dish with 30 mLs of modified E3 solution.
- Prepare an injection tray for the embryos. Our preferred injection mold is made of 1.5% agarose/E3, in which triangular depressions are formed such that the embryo can be positioned inside for injection. To make the injection mold, we float a plastic mold with wedge shaped protrusions (described in detail previously11) in a Petri dish filled with a base of 1.5% agarose.
- Prepare glass microinjection needles by pulling capillary tubes with a needle puller, as described in the Zebrafish Book10. In brief, pull the glass capillary tubes then use fine metal forceps to cut the tip of the microinjection needle to make a tip with a sharp point while viewing the needle tip under a stereomicroscope. Store the cut needles covered in a petri dish and positioned on modeling clay to protect the cut edge and prevent dust accumulation.
- Prepare the injection solution to label the proximal tubule. Dissolve 40 kD dextran-fluorescein conjugate (Invitrogen, D1845) at a concentration of 1 mg/mL in distilled water.
- To anesthetize the fish, add 5 mLs of 0.2% Tricaine pH 7.0 to the petri dish containing the zebrafish larvae in 30 mLs of E3. The fish are anesthetized when they no longer exhibit the touch response. It is vital that the fish are completely anesthetized or they will twitch upon attempting to inject them.
- Transfer the anesthetized fish to an injection mold tray using a plastic transfer pipet. Position the animal with its head in the deepest part of the triangular depression well, and on its side such that the trunk is resting along the angled side of the well.
- Perform intramuscular microinjection. Load a cut needle with 2-3 μL of dextran fluorescein, and inject the animal in a trunk somite with approximately 1 nL of solution. Aim for the upper portion of the somite, and avoid the yolk sac extension. This ensures that the nephron tubules are not mechanically disrupted by the microinjection process.
- Transfer injected zebrafish to a clean petri dish, and add modified E3 solution. Rinse the animals 3 times with fresh E3 to remove all traces of Tricaine. Cover the lid of the petri dish with aluminum foil to provide light-protection for the dextran, and return the animals to the 28°C incubator overnight.
2. Targeted laser ablation of proximal tubule cells
For this procedure, we use a stereomicroscope with epifluorescence to score animals with dextran fluorescein brightly-labeled proximal tubules and select these for laser ablation. Then, we use a pulsed micropoint laser system (Photonic Instruments, Inc) that has been attached to a compound microscope (Nikon Eclipse 80i with epifluorescent attachment), and calibrated for use as per manufacturer instructions, to perform nephron cell ablation. We have had the most success in cell ablation by using a FITC filter that enables the researcher to view the fluorescent tubule regions under brightfield lighting.
In advance of this procedure, prepare immobilization media for the ablation by dissolving 1.5% methylcellulose in 0.02% tricaine/E3. Store aliquots of methylcellulose for immediate use at room temperature, or at 4°C for long-term storage.
- Anesthetize the 72 hpf zebrafish by adding 5 mLs of 0.2% Tricaine pH 7.0 to the petri dish containing the zebrafish larvae in 30 mLs of E3. The fish are anesthetized when they no longer exhibit the touch response. It is vital that the fish are completely anesthetized or they will twitch upon attempting to perform the laser ablation.
- Examine the injected animals under the appropriate fluorescent setting and select animals in which you can easily visualize the proximal tubules.
- Transfer selected embryos to a separate dish containing tricaine. Animals can be anesthetized for up to 30 minutes before performing cell ablation. If you score >15 animals, return these to a separate dish containing fresh modified E3 while awaiting ablation as each cell ablation takes several minutes.
- To perform the ablation, transfer one anesthetized zebrafish to a small petri dish containing 1.5% methylcellulose/0.02% tricaine for 2 minutes to wash the embryo in immobilization media.
- Next, transfer the embryo to a glass depression slide containing a drop of 1.5% methylcellulose/0.02% tricaine. Gently position the animal with a fine probe such that its ventral side faces the slide and dorsal side is facing up.
- Place the slide on the compund microscope stage, and focus on the dorsal side of the animal. Perform ablation of the nephron cells under focus by depressing the foot pedal. You will see dispersal of the fluorescence as the renal epithelial cells are ablated (Figure 1A).
- Gently transfer the ablated animal to a well in a 12 well-dish for subsequent culturing. Rinse the E3 2-3 times to wash away the methylcellulose.
- After injection, raise the animal to the desired timepoint and perform desired analysis as per established histological and molecular techniques for young zebrafish embryos (Figure 2).
3. Representative results:
The data that are shown in Figure 1 indicate the ablation timecourse. After intramuscular injection with dextran-conjugates, the proximal tubule is preferentially labeled. Injection of dextran-FITC was used to label the kidney for laser ablation, and ablated animals were reinjected with dextran-rhodamine at one additional timepoint following ablation to reimage the proximal tubule population. Expression analysis techniques like in situ hybridization can be used to analyze changes in fixed samples at single timepoints following cell ablation, as shown in Figure 2.
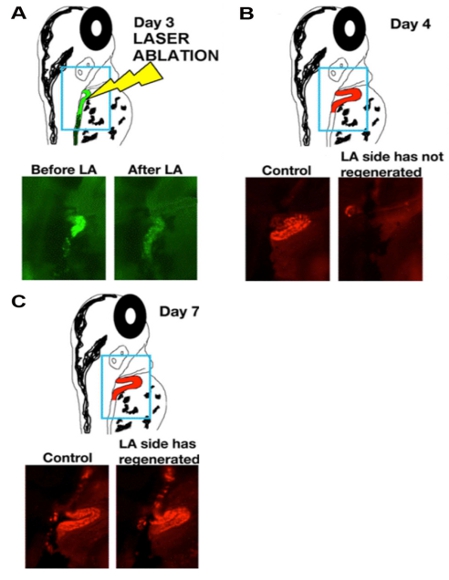
Figure 1: Laser ablation procedure is followed by rapid regeneration of the tubule epithelium. (A-C) Timecourse of cellular changes during and subsequent to laser ablation of zebrafish larva proximal tubules at time of ablation (day 3, panel A), or one or three days post ablation (day 4, panel B; day 7, panel C). (Top) schematics show position of the nephron, with dextran-FITC (green) and dextran-rhodamine (red), and (bottom) live images of control and laser-ablated (LA) nephrons.
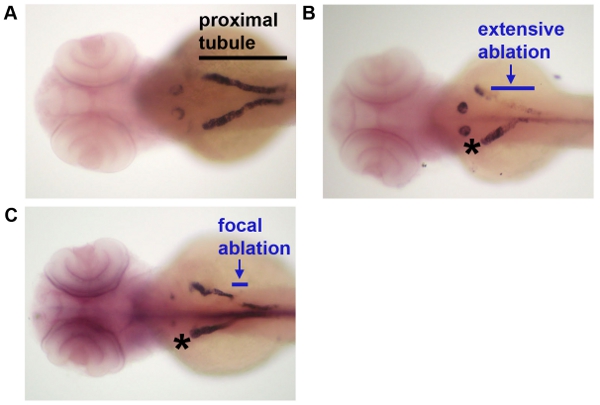
Figure 2: Analysis of gene expression following laser ablation. Following laser ablation at day 3, embryos were fixed and processed by whole mount in situ hybridization to detect transcripts for slc20a1a, which mark the proximal convoluted tubule segment of the nephron. (A) A control embryo that was not subjected to laser ablation, (B) an embryo in which a large stretch of tubule epithelium was ablated, and (C) an embryo in which a short interval of tubule epithelium was ablated. Black line indicates the extent of the proximal tubule in A to provide reference, blue lines indicate the extent of laser ablation in panels B-C, and * indicates the control (non-ablated) contralateral nephron in panels B-C.
