Embryo handling devices are pictured in Figure 1 to assist in visualizing the “home-made” devices for manipulation in the above Protocols. Results seen in Figure 2 illustrate the robust effect of rearing embryos at 18 °C on their ability to be permeabilized by EPS at late stages of development. This condition is applied in the protocol step 2.1. Efficacy of the CY5 carboxylic acid dye to reveal the various levels of permeability typically seen in EPS treated embryos is seen in Figure 3. The developmental dynamics of the dye distribution in the yolk is also seen in Figure 3, revealing a criterion used to assess viability, as described in Protocol step 4.2. The utility of the CY5 dye in determining embryo permeabilization subsequent to toxin treatment, formaldehyde fixation and immunostaining is illustrated by the result in Figure 4.

Figure 1. Embryo handling devices for the EPS method. The flat-bottomed basket is used in dechorination and EPS exposure steps (A,A'). The development basket is used for longer developmental exposures of permeabilized embryo (B,B'). The slide chamber is used for shorter developmental exposures and higher resolution imaging of live embryos (C,C'. See text for further description).
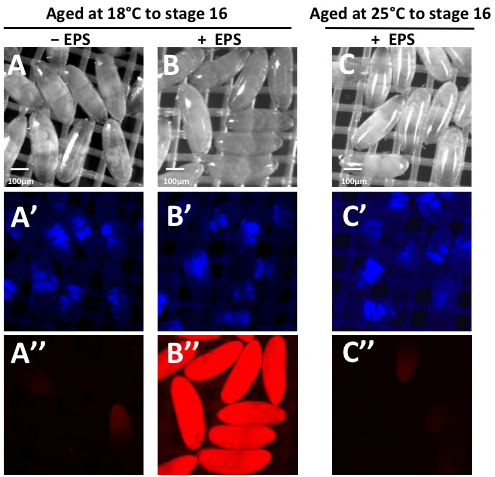
Figure 2. Effect of aging at 18 °C on EPS efficacy in late stage embryos. Embryos were collected for two hours followed by aging at 18 °C for 20 hr (Panels A-A”, B-B”) or at 25 °C for 10 hr (Panel C-C”). Embryos at 18 °C were then dechorionated and divided into two samples. The first sample was treated directly with 1 mM Rhodamine B dye in MBIM-T for 5 min, washed and visualized under brightfield and blue and red fluorescence channels (Panel A-A”). The second sample was treated with EPS (1:10 in MBIM for 1 min), washed and then treated with 1 mM Rhodamine B for 5 min and washed before visualization (Panel B-B”). Embryos raised at 25 °C were dechorionated and treated directly with EPS (1:10 in MBIM for 1 min), washed and then treated with 1 mM Rhodamine B for 5 min before visualization (Panel C-C”). Embryos were determined to be at stage 14 by the folds in the gut revealed by yolk autofluorescence in the blue channel (Panel A’, B’, C’). Embryos raised at 18 °C are impermeable prior to EPS treatment as seen by absence of Rhodamine B uptake (Panel A”). EPS treatment of 18 °C embryos yields a high degree of permeability as seen by Rhodamine B uptake (Panel B”). Embryos raised at 25 °C remain impermeable even with EPS treatment as seen by exclusion of Rhodamine B (Panel C”).
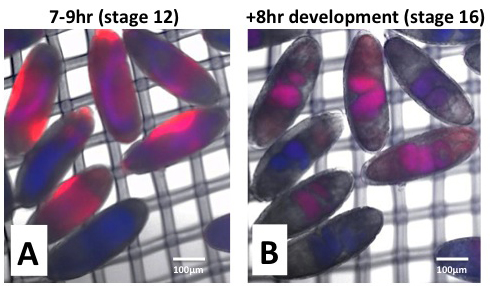
Figure 3. Incorporation of CY5 in permeable and viable embryos. Embryos were collected at 25 °C for 2 hr and aged for 14 hr at 18 °C (equivalent to 7-9 hr embryos at 25 °C, stage 12). After dechorination, EPS treatment was done (1:40 in MBIM for 1 min) followed by incubation in CY5 dye (50 µM in MBIM-T for 15 min). Embryos were washed three times in MBIM-T and transferred to development basket with MBIM in the reservoir. Development was allowed to proceed for 8 hr at room temperature. Uptake of CY5 (Red) is imaged in the far-red channel immediately after dye treatment and washing (Panel A) and after 8 hr development (Panel B). Distribution of the yolk is seen by autofluorescence in the blue channel. Dye uptake, hence permeability, is seen to vary from embryo to embryo. CY5 dye (red) is seen to localize to the yolk (blue), which becomes concentrated to the lumen of the gut at stage 16 (purple, Panel B).
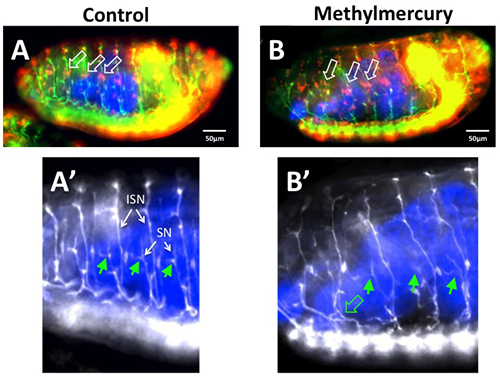
Figure 4. Determination of permeabilization and methylmercury effects in fixed and immunostained embryos. Embryos were collected at 25 °C for 2 hr and aged for 14 hr at 18 °C (equivalent to 7-9 hr embryos at 25 °C, stage 12). After dechorination, EPS treatment was done (1:40 in MBIM for 1 min) followed by incubation in CY5 dye (50 µM in MBIM-T for 15 min) together with methylmercury (50 µM MeHg, Panel B) or DMSO solvent control (0.1% final concentration, Panel A). Embryos were washed with MBIM-T and placed in a development basket with MBIM:M3 medium in the reservoir and aged for an additional 8 hr at room temperature. Embryos were then fixed in a two-phase 4% paraformaldehyde-heptane preparation by a standard protocol14. Staining was performed with anti-Fasciclin II (green in A,B and white in A’,B’) to label motor neurons and anti-elav antibodies (red in A, B) to label all neuron cell bodies. CY5 dye is revealed by direct fluorescence, which requires extended exposure due to diminished fluorescence intensity due to fixation (CY5 is pseudo-colored blue in all panels). The effects of MeHg are seen in the irregular patterning and clustering of the lateral chordotonal neuron cell bodies (elav-positive, labeled in red and denoted with white arrows in B versus A). In addition, a characteristic branching of the segmental (SN) (solid green arrows in A’) is seen to be highly variable with MeHg exposure (solid green arrows in B”) consistent with previously reported effects of MeHg on the embryo15. Projection of the intersegmental and segmental nerves at their roots are seen to be displaced posteriorly with MeHg exposure (open green arrow in B’). Note: Methylmercury is a potent neurotoxin. Care should be taken to wear gloves and eye protection when handling. Disposal should be done through an institutional environmental safety facility and service.
