The recording of human cortical slice activity starts while perfusing the tissue with normal recording ACSF (as previously described)7,8. In this condition, when the tissue is well preserved during surgery and later on during tissue transfer from the hospital and slice preparation, one can observe spontaneous activity, in the form of small interictal-like events, detected from small clusters of MEA electrodes. Interictal-like discharges consisted in a field potential, usually biphasic, comprising an initial sharp negative deflection, reaching tens of microvolts, followed by a longer opposite wave lasting several tens of milliseconds. Fast multi-unit activities are usually embedded in the field potential mainly in the initial part. A representative example of interictal-like activity is illustrated in Figure 3Aii, where the trace recorded by a MEA electrode is shown.
To record epileptic seizures from in vitro human cortical slices, it is necessary to perfuse them with an “epileptogenic” ACSF, in which Mg2+ is absent and K+ is raised to 6 mM, to boost tissue excitability1 (the increase of K+ from 3 to 6 mM alone is not enough to induce seizures; data not shown). Once the perfusion with 0 Mg2+ 6 mM K+ ACSF has started, the activity of cortical slices gradually increases and the first seizure appears within 15 to 20 min (15.4 ± 1.7 min, n = 10, from 5 patients; Figure 3 Ai). We evoked ictal-like activity, as shown in Figure 3, in 75% of patients tested and in 66.7% of all recorded slices.
Epileptic activities are recorded from adjacent electrodes of the MEA chip, and comparison of the onset dynamics of field potential events allows studying their site of genesis and propagation. A representative seizure recorded from a slice of human cerebral cortex with the MEA system is shown in Figure 3B (a representative trace of a seizure recorded by a single MEA electrode is shown at higher temporal resolution in Figure 3 Aiii). Note that the seizure is recorded from almost all the electrodes of the MEA chip, thus indicating that it is able to propagate to a large cortical area. Furthermore, looking at the single traces recorded from each electrode, it is possible to observe the progressive propagation of the seizure across the tissue: indeed, it starts first in the cortical area covering the right half of the electrode array, and it gradually spreads to the area covering the left half (i. e., compare traces from electrode L6 and B6; Figure 3B).
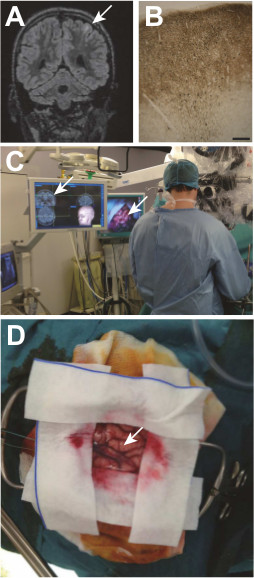
Figure 1: A cortical dysplasia and its surgical removal. A Taylor type focal cortical dysplasia (white arrow) embedded in a left frontal lobe sulcus is visualized on the preoperative MRI (A), and later confirmed by histological examination (B) MAP kinase labeling; scale bar: 200 μm) showing a cortical dyslamination, abnormal neurons and a track on neurons with an incomplete migration in the lower white matter. During surgery, the dysplasia is localized according to the MRI data with a neuronavigation system (C). Visualization of the gyrus containing the dysplasia during surgery (D).
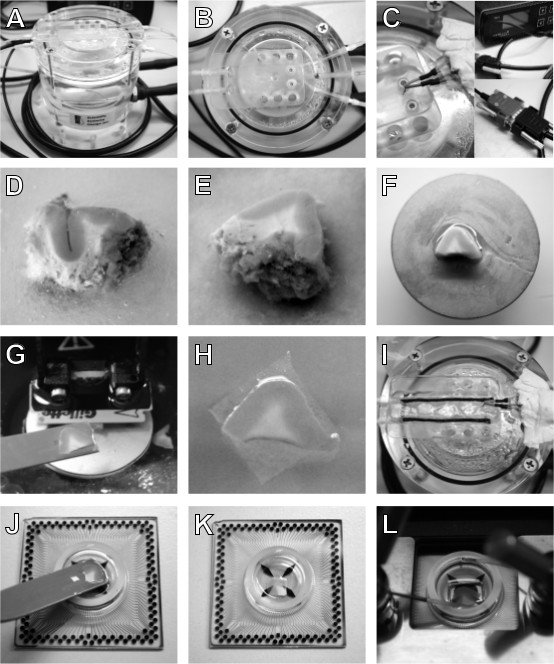
Figure 2: Equipment and procedure for human cortical slice preparation. An interface chamber is used to maintain human cortical slices ex vivo (A); the slices rest on a sheet of lens tissue in the flat area in the upper part of the chamber (B), where a temperature sensor (C, left), connected to a temperature controller (C, right, up) through a double-ended cable (C, right, down), is placed to maintain 37 °C throughout slice incubation time. Once obtained, place the resected block of tissue in a petri dish filled with ice-cold sucrose-based ACSF (D) and remove blood clots, vessels and meninges, to facilitate the slicing. If the tissue block is bigger than MEA chambers, isolate a piece of appropriate dimensions with a blade (E) and glue it on the vibratome specimen plate (F). Cut 400 µm thick slices and with the help of a spatula (G) transfer them on pieces of lens tissue (H) and incubate them in the interface chamber (I). Before recording, remove the lens tissue by submerging the slice in a petri dish filled with ACSF and transfer it to an ACSF filled MEA chip with a spatula (J). Remove the solution to let the slice adhere on the MEA (K) and keep it in position by using a platinum anchor. Place the MEA in the amplifier (L), start perfusion and recording.
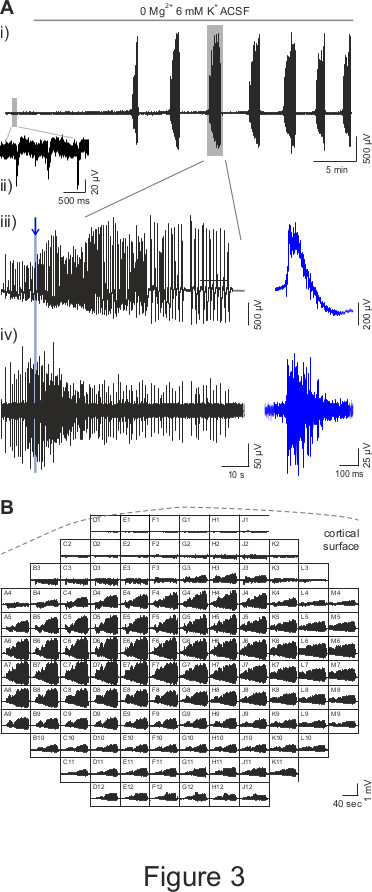
Figure 3: MEA recordings of epileptic seizures in human cortical slices. (A) A representative trace of human cortical slice activity recorded by a MEA electrode is shown in panel i. In presence of normal ACSF, it is possible to observe interictal-like spontaneous activity (small grey rectangle, zoomed in panel ii); then, when the perfusion with 0 Mg2+ 6 mM K+ ACSF has started, the first seizure appears after a ~15 min delay and is followed by other events at 2-3 min interval. Panel iii shows the seizure in the big grey rectangle in panel i with a zoomed scale. The blue trace illustrates a zoom of the activity, as indicated by the blue line and arrow. Panel iv) shows the data illustrated in panel iii) high pass filtered at 250 Hz, which reveal multi-unit activity when zoomed (blue trace). (B) Representative recording of a seizure in presence of 0 Mg2+ 6 mM K+ ACSF from all MEA electrodes. Each square represents an electrode of 12 x 12 MEA array and shows an 80 sec time window; the dotted line indicates the position of the cortical surface relatively to MEA array.
