To assess the difference between fTAT and dfTAT, HeLa cells are incubated for 1 hr with each peptide to determine the difference in their cellular localization. The internalization of the two CPP’s was assessed using fluorescence microscopy. Figure 2 shows that fTAT (20 µM) localizes in a punctate distribution. This distribution is consistent with the peptide remaining entrapped inside endosomes. In contrast, the fluorescence signal of dfTAT (5 µM) displays a homogenous distribution throughout the cytosol and nucleus. Figure 3 shows that the cytosolic distribution of dfTAT is observed in a number of different cell lines including COLO 316, NIH 3T3, HaCaT, the difficult to transfect Neuro-2a, MCH58, the primary cell line HDF, DRG-F11 and NCL-H1299. The 20X images shows that a very high percentage (>80%) in a dish display a cytosolic distribution of dfTAT with no cellular toxicity (no Sytox blue nuclear staining).
To determine whether dfTAT-mediated endosomal leakage delivers large proteins into the cytosol of cells. EGFP (26 kDa) is chosen as a model protein. This is because its fluorescence can be used to detect its delivery into the cells by observing the localization of the green fluorescence (if correctly folded). To do this assay, EGFP and dTAT were incubated for 1 hr with cells, as shown in Figure 4, EGFP displayed a cytosolic and nuclear green fluorescent distribution similar to what is observed for dfTAT in more than 90% of cells without observable toxicity.
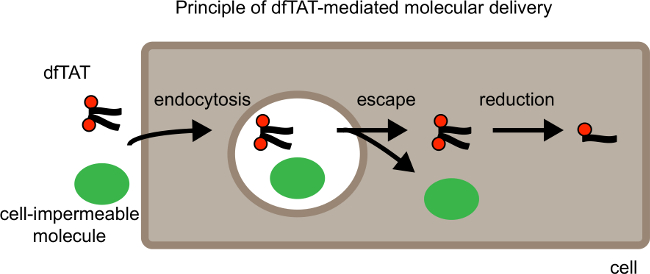
Figure 1. Schematic Showing Principle of dfTAT-mediated Molecular Delivery. From left to Right. Schematic shows dfTAT cellular delivery along with a cell impermeable cargo. First dfTAT induces endocytosis which results in the uptake of dfTAT along with the cargo into endocytic vesicles. In the second step dfTAT escapes from the endocytic vesicles which results in the release of dfTAT and the cell impermeable molecule into the cytosol of cells. The cytosol of mammalian cells is a reductive environment, therefore in the cytosol dfTAT is reduced to its monomer counterpart fTAT. Please click here to view a larger version of this figure.
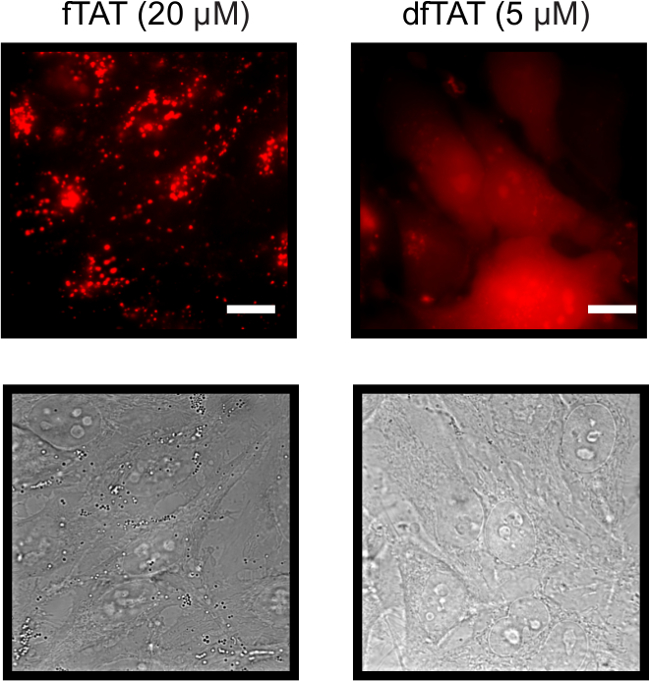
Figure 2. Fluorescence and Bright Field Images of Both HeLa cells Incubated with 20 µM fTAT (left panel) and 5 µM dfTAT (right panel) using a 100X Objective. fTAT monochrome images shows cells that exhibit a fluorescence punctate distribution while dfTAT images shows cells displaying a homogenous cytosolic and nuclear fluorescence distribution. Scale bar: 10 µm. Please click here to view a larger version of this figure.
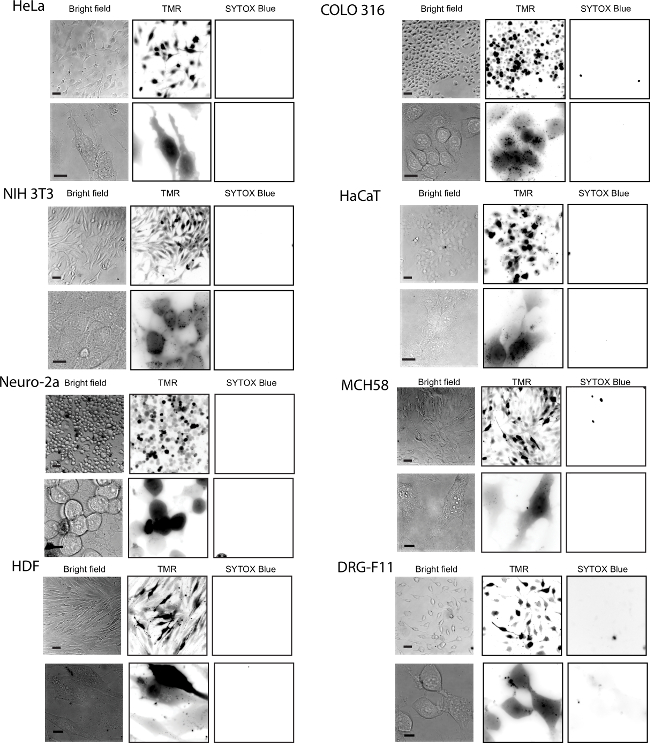
Figure 3. Efficient Delivery of dfTAT into Live Cells was Achieved in Multiple Cell Lines. The cell lines tested were the following: HeLa, NIH 3T3, COLO 316 and HaCaT, Neuro-2a, MCH58, HDF, DRG-F11 and NCL- H1299. Cells were incubated with 5 µM dfTAT for 1 hr followed by a washing step according to protocol and imaged. The fluorescence signal detected was in the cytosol and nucleus of cells (top panel: 20X objective, bottom panel: 100X objective). The cell-impermeable nuclear stain SYTOX Blue was used to assess cell viability after dfTAT treatment. Scale bars, 20X objective: 50 µm; 100X objective: 10 µm. Please click here to view a larger version of this figure.
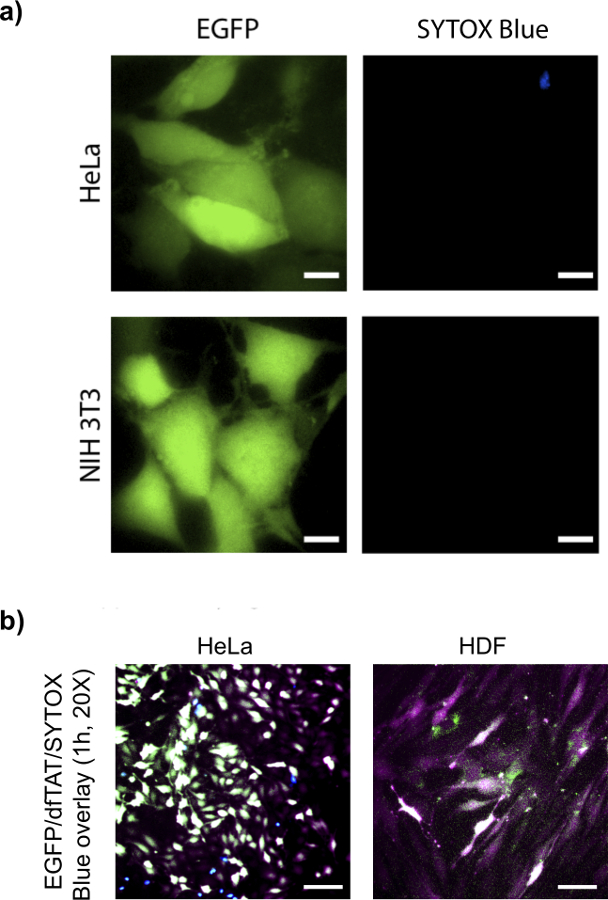
Figure 4. dfTAT Delivers Intact EGFP into Different Cell Lines. (A) HeLa (top panel) and NIH 3T3 (bottom panel) cells were incubated with EGFP (10 µM) and dfTAT (5 µM) for 1 hr and following step were performed according to protocol. Images show a homogenous cytosolic fluorescence distribution of EGFP in both cell lines. Scale bars, 10 µm. (B) HeLa and primary HDF cells were incubated with EGFP (10 µM) and dfTAT (5 µM) according to protocol. 20X images show a homogenous cytosolic fluorescence distribution of EGFP and dfTAT in HeLa and primary HDF cells. Overlay (pseudocolor: white) indicate presence of dfTAT (pseudocolor: purple), EGFP (pseudocolor: green) in the cytosolic space of both HeLa and HDF cells. Sytox blue (shown in blue) was used to asses for cell viability. Scale bars: 50 µm. Please click here to view a larger version of this figure.
