We first investigated the optimal concentration of PVP using Arabidopsis mature seeds. Total RNA was isolated from approximately 1,000 seeds according to the protocol described above using cell lysis buffer containing 0%, 0.25%, 0.5%, 1.0% or 2.0% PVP. After homogenization and centrifugation, the supernatant was collected while avoiding the oil layer and seed debris (Figure 1A).
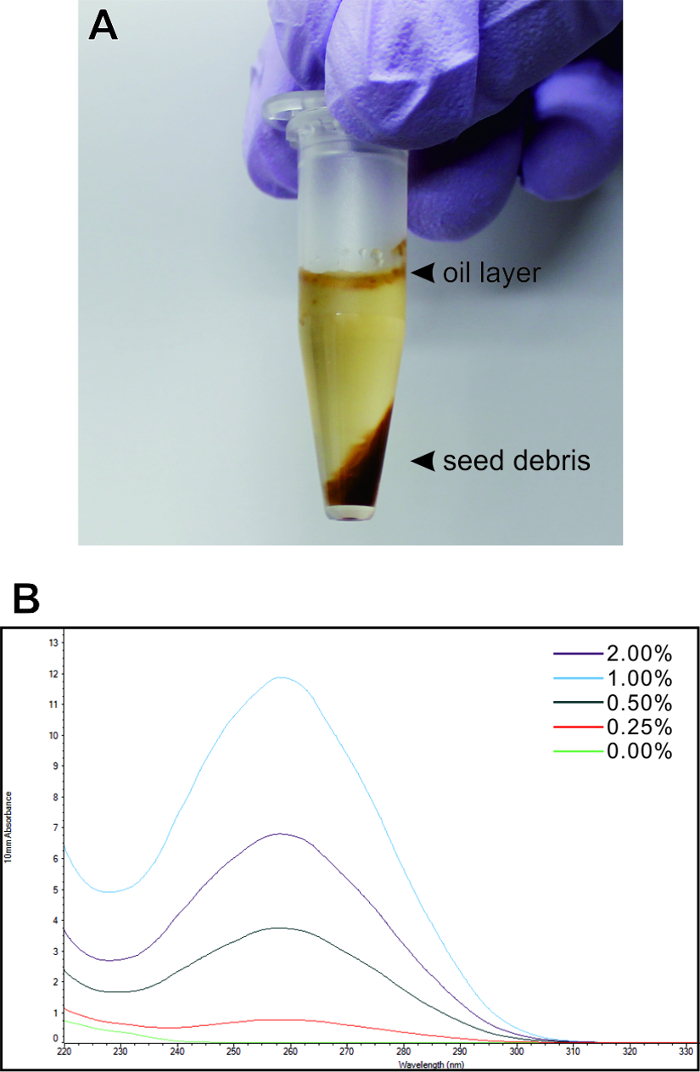
Figure 1: Estimation of the optimal polyvinylpyrrolidone concentration in the cell lysis buffer. Total RNA was extracted with the cell lysis buffer containing different concentrations of PVP. (A) A photograph of the seeds after homogenization and centrifugation. (B) The graphs of wavelength absorbance for each sample are indicated. Please click here to view a larger version of this figure.
The quantities and purity of the isolated RNAs were estimated from the graph of wavelength absorbance (Figure 1B), which showed that 1.0% PVP was the most effective concentration of isolating large amounts of purified RNA from seeds.
Developing seeds were isolated from Arabidopsis thaliana fruits on pre-chilled aluminum plates on ice under a stereomicroscope (Figures 2A, 2B). The seeds were homogenized with a stainless steel pestle and motor-grinder in a tube on an aluminum rack on ice (Figures 2C, 2D).

Figure 2: Isolation and homogenization of developing seeds of Arabidopsis thaliana. Developing seeds are isolated from fruits on a cold aluminum plate under a stereomicroscope (A). Carpels are peeled from the pedicle (B), and developing seeds are collected. The seeds are homogenized in lysis buffer containing 1% (w/v) polyvinylpyrrolidone using a stainless steel pestle (C) in tubes in an aluminum rack chilled on ice (D). Please click here to view a larger version of this figure.
Total RNA was isolated from the seeds at 4, 8, and 12 days after flowering (hereafter referred to as DAF) using our newly developed method or the conventional method. The concentrations, A260/A280 ratios, and A260/A230 ratios of RNA isolated from 200 seeds are shown in Table 1.
| Method | DAF | RNA concentration | A260/A280 | A260/A230 |
| Conventional | 4 | 72.4 ± 7.4 | 1.90 ± 0.02 | 2.27 ± 0.06 |
| 8 | 10.8 ± 3.0 | 1.50 ± 0.08 | 1.40 ± 0.17 | |
| 12 | 4.9 ± 1.7 | 1.57 ± 0.10 | 0.76 ± 0.18 | |
| Nuevo | 4 | 63.7 ± 2.6 | 2.10 ± 0.08 | 2.29 ± 0.03 |
| 8 | 46.3 ± 4.9 | 2.15 ± 0.05 | 2.23 ± 0.09 | |
| 12 | 41.7 ± 3.1 | 2.09 ± 0.04 | 2.17 ± 0.07 |
Table 1: Concentrations, A260/A280 ratios, and A260/A230 ratios of RNA isolated from Arabidopsis seeds. Total RNA was isolated from approximately 200 seeds (from 190 to 206 seeds) at 4, 8, and 12 DAF using the conventional method or our newly developed method. The RNA concentrations, A260/A280 ratios, and A260/A230 ratios were measured. Values represent mean ± SD of three independent experiments.
The results show that our method enabled us to isolate highly purified RNA from 8 and 12 DAF seeds. The total RNA was reverse-transcribed to cDNA, and the cDNA solutions were diluted 100-fold. The copy numbers of WRI1 (WRINKLED 1: AT3G54320), SDP1 (SUGAR DEPENDENT1: AT5G04040)7, and EIF3K (eukaryotic translation initiation factor 3K: AT4G33250) as an internal control8 were measured, and the relative amounts of the transcripts were estimated by quantitative real-time PCR analyses (Figure 3).
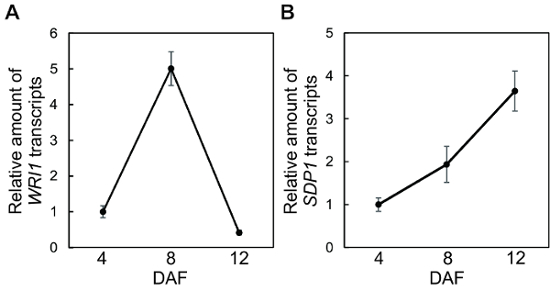
Figure 3: Quantification of low level transcripts in developing seeds. Total RNA was extracted from 4, 8, and 12 DAF seeds using our method, and cDNA solutions were prepared. The relative amounts of WRI1 (A) and SDP1 (B) transcripts were measured by quantitative real-time PCR. EIF3K was used as an internal control. Values represent mean SD of three independent experiments. DAF; days after flowering. Please click here to view a larger version of this figure.
Although the amounts of WRI1 and SDP1 transcripts are relatively low in developing seeds, it was possible to detect expression changes between seeds at different developmental stages. To verify that our method can be utilized in other oilseeds, total RNA was isolated from mature seeds of Brassica napus and Glycine max. The RNA concentrations, A260/A280 ratios, and A260/A230 ratios are shown in Table 2.
| Method | Plant | RNA concentration | A260/A280 | A260/A230 |
| Conventional | B. napus | 7.7 ± 0.8 | 1.5 ± 0.12 | 0.75 ± 0.11 |
| G. max | 9.7 ± 2.3 | 1.61 ± 0.06 | 0.69 ± 0.09 | |
| New | B. napus | 143.2 ± 19.0 | 2.17 ± 0.03 | 2.23 ± 0.13 |
| G. max | 152.3 ± 4.4 | 2.17 ± 0.02 | 2.30 ± 0.02 |
Table 2: Concentrations, A260/A280 ratios, and A260/A230 ratios of RNA isolated from mature seeds of Brassica napus and Glycine max. Total RNA was isolated from approximately 100 mg of Brassica napus and Glycine max seeds using the conventional method or our newly developed method. The seeds were previously crushed and roughly homogenized with a mortar and pestle chilled in liquid nitrogen. The RNA concentrations, A260/A280 ratios, and A260/A230 ratios were measured. Values represent mean ± SD of three independent experiments.
