Flies of the w1118 genotype are used to demonstrate how the assay is performed. The w1118 mutants are commonly used to generate transgenic lines and to control the genetic background of transgenes marked with the white gene. Normally, for behavioral experiments, all transgenic lines are backcrossed for five generations to the same w1118 stock, which is used as an experimental control. We show different experiments: a comparison of evaporation loss for our modified setup, a short-term food choice experiment, a long-term food intake experiment, and an experiment on different sucrose dilutions.
Evaporation plays a critical role in the performance of the CAFE assay. We included additional approaches to our assay to decrease evaporation: i) the central sponge bung is refilled with water every 24 h; ii) additional water filled vials within the transport box and iii) the use of a cover for the box to create a humidity enclosure (see 4.6). Comparing the evaporation between a setup without and with above mentioned devices, a significant reduction in evaporation is seen. Even the effect of higher volatility of an ethanol containing solution is not detectable using the new setup.
In a two-choice food experiment a group of 20 flies can feed for 3 h. In natural environments, fruit flies feed preferentially on fermenting fruits with alcohol22, and it has been shown, using a similar setup, that flies prefer yeast-sucrose solutions with ethanol over yeast-sucrose solutions without ethanol23. Here, two food choices are offered, a 0.1 M sucrose solution labeled with red food color and a 0.1 M sucrose solution with 15% EtOH labeled with blue food color (Figure 1A, C). Visual examination of the abdomen indicates that the flies feed on both solutions (Figure 1D). Food consumption per fly is significantly greater (nearly 2-fold) for the sucrose solution containing EtOH (Figure 3A).
In a following experiment, a long-term study, a group of eight flies has access to similar food sources for 4 days, and flies consume more of the ethanol-containing food on each day (Figure 3B). The preference index for ethanol ([Suc + EtOH] – [Suc]/total consumption) remains constant over this period (average = 0.29, Table 4). The observed ethanol preference is consistent with several other publications and shows that flies can distinguish between different food sources24,25,26. The observed ethanol attraction might be a result of the different caloric contents of the offered solutions and of the rewarding properties of ethanol24. The assay can also be used to measure negative effects of food supplements. Ja and colleagues showed in the first publication of this method that application of paraquat (an oxidizing drug) decreases food consumption10.
In the next experiment, the difference in food intake between the sexes is shown. Metabolic requirements differ between male and female D. melanogaster. For example, while male flies prefer carbohydrate-rich food, during egg production, a phase that requires increased protein biosynthesis, females prefer protein-rich diets over carbohydrate-rich diets27. Mated male and female flies were used in this experiment. To analyze differences in food intake between 20 male and 20 female flies within a 3 h feeding interval, a CAFE assay is performed using a sucrose concentration series. Five capillaries were provided, with solutions ranging from 10–3 to 2 M sucrose, and consumption of each solution was measured (Figure 4A). Results showed that both sexes preferred high-concentration sucrose solutions as a food source (Figure 4A). However, females consumed significantly more of the two lowest-concentration sucrose solutions compared to males (P < 0.05); on the other hand, males consumed significantly more of the higher-concentration solutions (P < 0.001). Note that these data did not account for differences in body size. Female D. melanogaster are usually larger and heavier than males (Table 1). When food consumption is normalized to fly mass, differences between males and females in consumption of low-sucrose solutions are no longer significant. In summary, males consume more sucrose solution than mated females, consistent with previous data, reflecting possible different metabolic demands, nutrient preferences or simple differences in the ability to feed on the capillaries between the two sexes.

Figure 1: The Drosophila melanogaster CApillary FEeder Assay. A) The feeding assay with flies. Moistened filter paper provides water at the bottom of the vial. Four capillaries are provided during the experiment (red- and blue-colored food in opposite capillaries). Note that the capillaries are secured in position by a second pipette tip, and unused positions are closed using pipette tips. A foam plug in the center of the lid allows air exchange. B) Detailed view of the lid. Cut pipette tips (2 – 20 µL, red borders) are inserted into the conical openings of unused positions, and a second pipette tip is inserted into the cut tip to close the hole. The cut pipette tips are used to control placement of the microcapillaries, and uncut tips are used to hold the capillaries tight. C) A D. melanogaster fly feeds on a capillary. D) After feeding, food color is clearly visible in the fly abdomen. E) A digital caliper is used to measure the distance between markbeginning and markend of the meniscus. The data are transferred directly to an Excel spreadsheet via USB. Please click here to view a larger version of this figure.
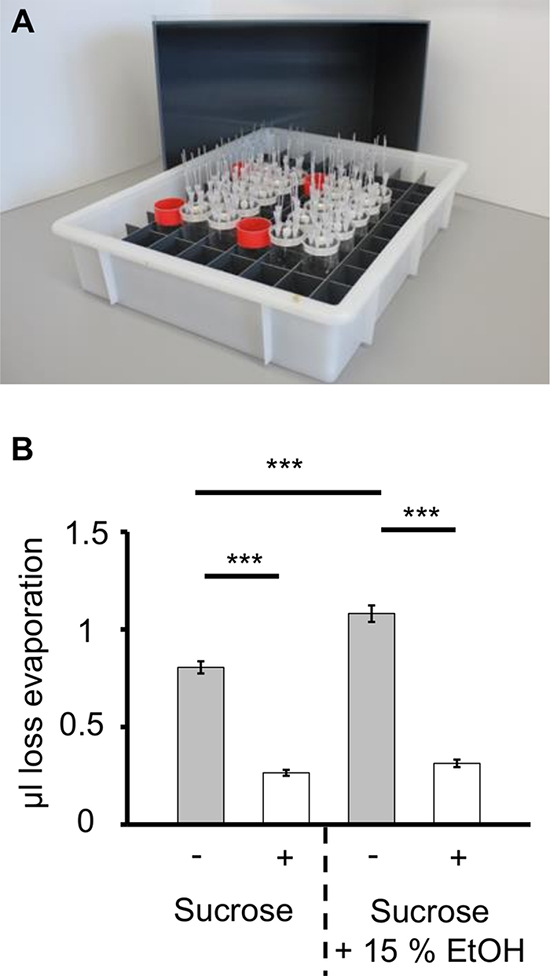
Figure 2: Influence of Evaporation in the Capillary Feeder Assay. A) Multiple CAFE assay placed inside a plastic box with a gridded inlay. For controlling the humidity during the experiment four water filled vials (red rims) are placed inside the grid. The evaporation controls are placed in direct proximity to these vials. A cover for the whole setup is shown in the background. B) Comparison of the volume loss through evaporation. The mean value for evaporation over 4 days is shown. Humidity is controlled by (i) applying water to the central sponge bung (24 h interval); (ii) adding four water filled vials into the grid; and (iii) using a plastic cover for the whole setup. The evaporation is significantly lower if humidity is controlled for both solutions tested (***P ≤ 0.001; N = 48). No differences in volatility between EtOH containing and non-containing sucrose solution is detectable with the humidity devices used. Please click here to view a larger version of this figure.
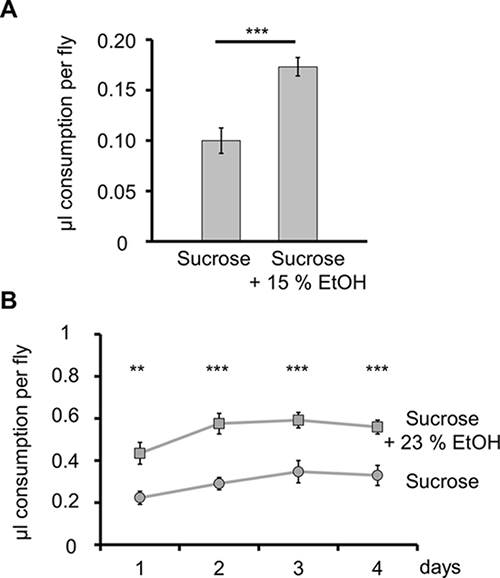
Figure 3: Preference for Ethanol (EtOH) Containing Sucrose over Sucrose Solution. A) Food consumption for male w1118 flies is shown. Males consume significantly more of a 15% EtOH containing sucrose solution than of a plain sucrose solution. ***P ≤ 0.001; N = 27. B) Flies significantly prefer a sucrose solution containing 23% EtOH during a 4-day trial. ***P ≤ 0.001; **P ≤ 0.01; N = 16. Please click here to view a larger version of this figure.
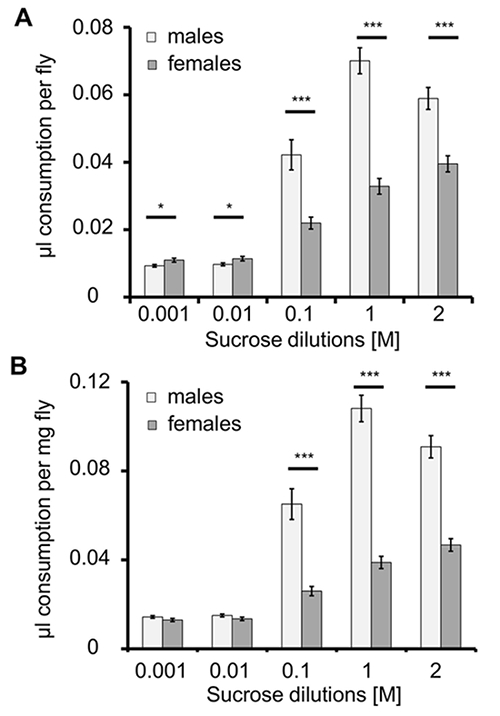
Figure 4: Consumption (µL/fly and µL/mg fly) of Different Sucrose Concentrations by Male and Female w1118 Flies. A) The consumption of different concentrations of sucrose solutions differs significantly between males and females. Female flies consume more at lower sucrose concentrations, and male flies consume more at higher concentrations. *P < 0.05; ***P < 0.001; N = 27 trials with 20 males each, N = 30 trials with 20 females each). B) Food uptake on a mass basis. A significant increase in consumption occurs between male and female flies for the 0.1 to 2 M sucrose solutions when normalized to fly mass. ***P ≤ 0.001; N = 27 males, N = 30 females. Please click here to view a larger version of this figure.
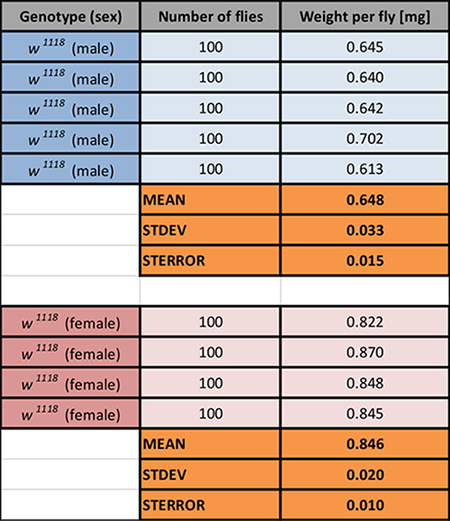
Table 1: Body Weight of Male and Female w1118 Flies. Four to five groups of 100 flies were measured, and body weight (mg/fly) was calculated. Mean values (with STDEV (standard deviation) and STERROR (standard error)) are shown. Mean values are used to normalize food consumption to fly mass (µL/mg fly). Please click here to download this spreadsheet.
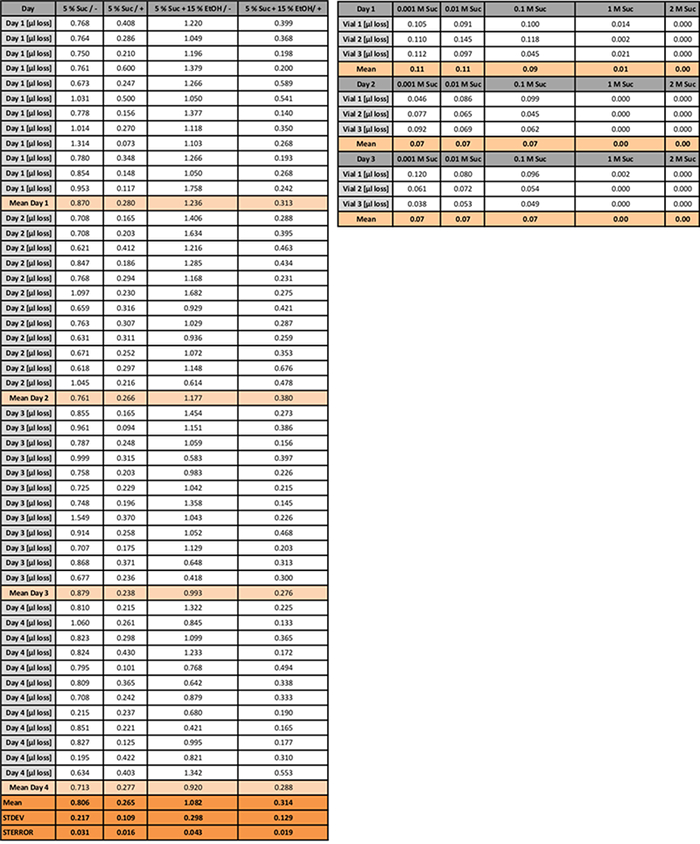
Table 2: Evaporation Loss (µL) in the CAFE Assay. The quantity of liquid lost through evaporation is shown for 4 days. Humidity is controlled (+) or not (-) as described in Figure 2. Evaporation data for two different solutions (sucrose and sucrose plus EtOH) are shown. Mean values are presented for each day and over the period (with STDEV and STERROR). The evaporation loss of the sucrose dilutions experiment is shown underneath separately (mean values). Please click here to download this spreadsheet.
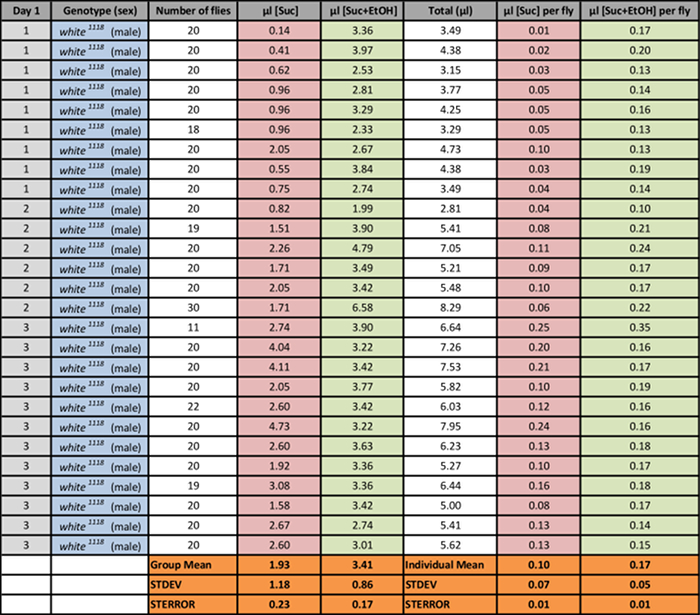
Table 3: Consumption of 0.1 M Sucrose with/ without 15% EtOH by Male w1118 Flies Fed for 3 h. Consumption of both solutions by groups of 20 flies was measured for 3 h on 3 days. Consumption values for fly groups are divided by the number of tested flies to estimate microliter uptake per fly after subtracting evaporative loss. Mean values (with STDEV and STERROR) are shown. Please click here to download this spreadsheet.
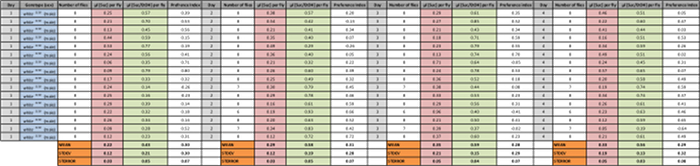
Table 4: Consumption of 0.1 M Sucrose With and Without 23% EtOH for Four Days by Male w1118 Flies. Consumption of both solutions by groups of 8 flies was measured for 24 h for 4 days. Preference index for ethanol was calculated by using the following formula ([Suc + EtOH] – [Suc]/total consumption). Consumption values for fly groups are divided by the number of tested flies to estimate µL uptake per fly after subtracting evaporative loss. Mean values (with STDEV and STERROR) are shown for each day. Please click here to download this spreadsheet.
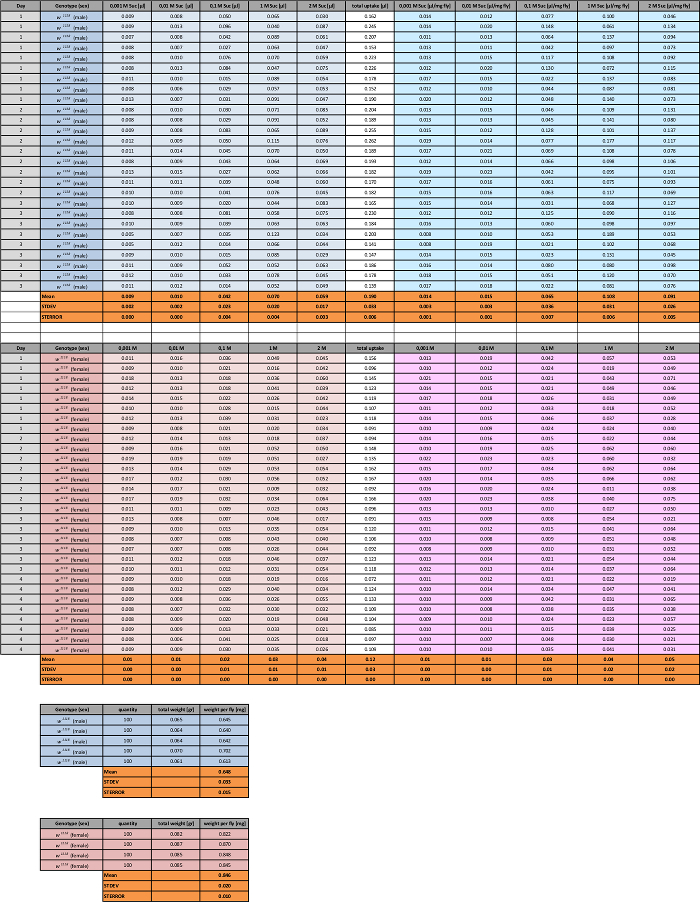
Table 5: Consumption of Five Concentrations of Sucrose by Male and Female w1118 Flies. Intake of each solution, and the value for the sum of sucrose intakes, is shown. Mean values for each concentration are given below each column (with STDEV and STERROR). To calculate intake based on fly mass (microliter uptake per milligram of fly), food consumption is divided by the average weight of male or female flies (from Table 1, shown to the right). Please click here to download this spreadsheet.
