The original findings of these experiments were described in depth previously7 and are reported herein as an example of results that can be obtained using the described protocols.
Maturation rate
Of the 32 COCs retrieved by OPU from dominant follicles, 28 (88%) were at the MII stage. Fourteen of the 58 COCs collected from follicles 5-25 mm in size were immediately snap-frozen as immature oocytes for the mRNA expression study. The remaining 44 were in vitro matured, and 37 reached the MII stage (85%). The efficiency of maturation was not significantly different in the two groups (Fisher's exact test P >0.05).
Aneuploidy rate
In order to study whether IVM can perturb the faithful chromosomal partitioning between the secondary oocyte and the first polar body, the number of chromosomes at the MII stage was counted, as it represents the result of chromosome segregation during meiosis I. As shown in Figure 1A, the anti-Aurora B phospho-Thr232 antibody specifically stained the centromeric region in horse oocytes, allowing one to count the number of chromosomes. In vitro-matured oocytes were significantly more affected by aneuploidy (5/11) compared to in vivo matured oocytes (0/12; Fisher's exact test, P <0.05; Figure 1B). Most of the aneuploid oocytes (4/5) were hyperploid; therefore, the aneuploidy rate does not result from artifacts in the preparation of the sample determining chromosomal loss, as happens with chromosomal spreads. Immunostaining of the centromeric region was also attempted with the anti-CENPA and anti-AURKB antibodies, but no signal was visualized in both cases (data not shown).
Spindle morphology and acetylation of H4K16
The pole-to-pole spindle length and the diameter of the spindle at the maximum width were measured, as illustrated in Figure 2. According to these measurements, in vitro-matured oocytes were significantly longer (19.89 ± 1.15 µm) and wider (17.28 ± 0.81 µm) compared to in vivo matured oocytes (14.57 ± 1.7 µm in length and 13.56 ± 0.91 µm in diameter; unpaired t-test, P <0.05). On the same samples, the global acetylation level of H4K16 was also measured, confirming that acetylation at H4K16 was significantly lower in IVM oocytes compared to their in vivo counterparts (Figure 2, red).
Expression of transcripts coding for histone acetyltransferases and deacetylases
Whether the expression of genes involved in the histone acetylation-deacetylation process was altered in in vitro-matured oocytes was investigated by q-PCR. Histone acetyl-transferase 1 (HAT1), K-acetyltransferase 8(KAT8), histone deacetylase 1 (HDAC1), and NAD-dependent protein deacetylase sirtuin 1 (SIRT1) were identified as putative targets. Significant differences in the expression of these transcripts in immature, in vitro matured, and in vivo matured oocytes were not observed, independent of the normalization used. Specifically, the results of the transcript expression analysis reported in Figure 3 were calculated against a standard curve using the SQ method and normalized for housekeeping glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Similarly, no differences were observed when the results where normalized for the spike in luciferase or when the ΔΔct method was used (not shown).
| Target | Fw primer | Rv primer | Amplicon size |
| GAPDH | ATCACCATCTTCCAGGAGCGAGA | GTCTTCTGGGTGGCAGTGATGG | 241 bp |
| HAT1 | CTGACATGAGTGATGCTGAACA | TAACGCGTCGGTAATCTTCC | 219 bp |
| HDAC1 | GAAGGCGGTCGCAAGAAT | CCAACTTGACCTCCTCCTTGA | 166 bp |
| KAT8 | ACTGGTCTTGGGTCCTGCTG | GGGCACTTTTGAGGTGTTCC | 198 bp |
| Luciferase | TCATTCTTCGCCAAAAGCACTCTG | AGCCCATATCCTTGTCGTATCCC | 148 bp |
| SIRT1 | GACTCGCAAAGGAGCAGATT | GGACTCTGGCATGTTCCACT | 169 bp |
Table 1: Primer Setails. The 5' > 3' sequence of the forward (Fw) and (Rv) primers and the expected amplicon size are given for each transcript analyzed: glyceraldehyde-3-phosphate dehydrogenase (GAPDH), histone acetyl-transferase 1 (HAT1), histone deacetylase 1 (HDAC1), K-acetyltransferase 8 (KAT8), luciferase, and NAD-dependent protein deacetylase sirtuin 1 (SIRT1). Reprinted with permission from reference7.
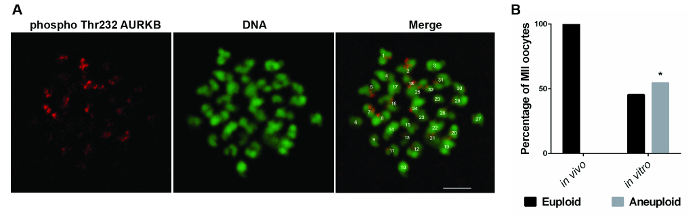
Figure 1: Aneuploidy Rate in In Vivo- and In Vitro-matured Horse Oocytes. (A) Representative images showing aurora B phospho-Thr232 (red) and DNA (green) staining of a MII-stage horse oocyte treated with monastrol. A euploid oocyte (32 chromosomes) is shown. Scale bar = 5 µm. (B) The bar graph represents the distribution of euploid and aneuploid MII-stage oocytes in in vivo (n = 12) and in vitro (n = 11) matured oocytes. *Indicates a significant difference in the aneuploidy rate (Fisher's exact test, P <0.05). Reprinted with permission from Reference 7. Please click here to view a larger version of this figure.
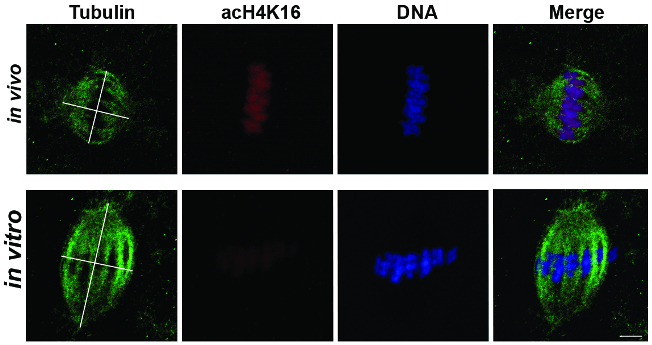
Figure 2: Spindle Measurement and H4K16 Acetylation in In Vivo- and In Vitro-matured Horse Oocytes. Representative images showing tubulin (green), acetylated H4K16 (red), and DNA (blue) in MII oocytes after in vivo (n = 4) or in vitro (n=14) maturation. The tubulin images show the axes used for spindle length and diameter measurements. Scale bar = 5 µm. Modified from reference7. Please click here to view a larger version of this figure.
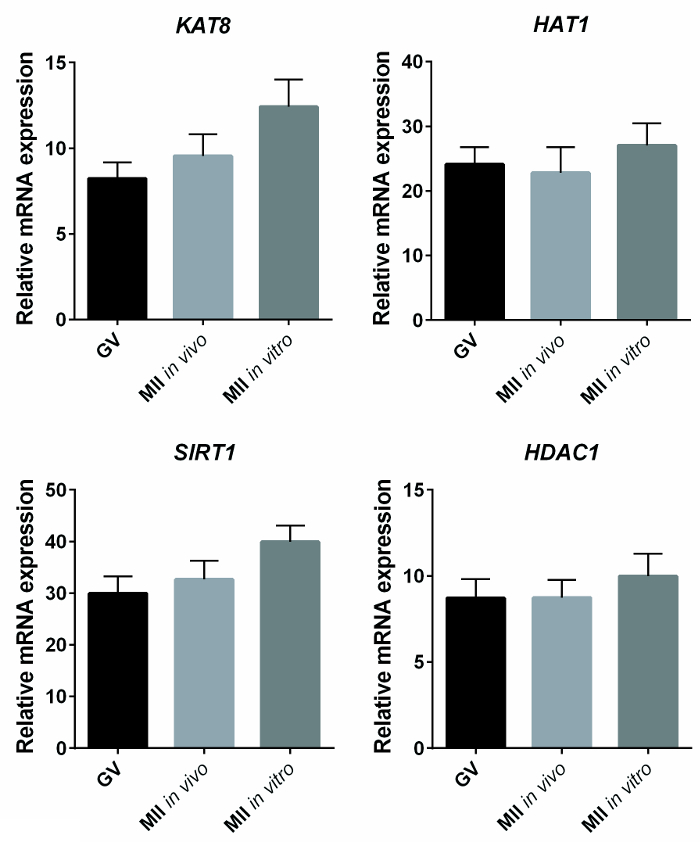
Figure 3: Expression of KAT8, SIRT1, HAT1, and HDAC1 in Immature, In Vivo, and In Vitro Matured Oocytes. The bar graphs represent the mean ± SEM of the relative mRNA expression of KAT8, SIRT1, HAT1, and HDAC1, expressed as the starting quantity (SQ) of the transcript of interest compared with the SQ of GAPDH, used as housekeeping. No statistical differences were observed between immature (GV, n = 14), in vivo (n = 14), and in vitro (n = 12) matured oocytes (one-way ANOVA, P >0.05). Reprinted with permission from reference7. Please click here to view a larger version of this figure.
