Lateral roots grow at physiological rates in imaging chambers.
Lateral root lengths of plants in imaging chambers were measured at hourly intervals (Figure 2A, left, Movie 1, n = 23) and growth rates were compared to those of lateral roots grown on standard Petri plates containing the same agar-solidified medium (Figure 2A, right, Movie 2, n = 23). Growth rates can vary considerably between lateral roots, with the length of the root being one determining factor owing to the increasingly longer zones of growth and proliferation13. Therefore, sets of lateral roots were selected to represent roots of different initial lengths (ranging from 50 µm to 1150 µm) in both the imaging chamber and the agar plate. Average lateral root length at the start of the experiment was similar in each set (439 and 442 µm for imaging chambers and plates, respectively). In the analyzed time interval of 27 h, lateral root growth was approximately linear both in imaging chambers and on plates, with an average growth rate of 52 µm/h (R2 = 0.997) on plates and 51 µm/h in imaging chambers (R2 = 0.993) (Figure 2B). Growth rates of individual lateral roots where highly variable both on plates and in imaging chambers (Figure 2C), ranging from 17 µm/h to 82 µm/h in imaging chambers and from 13 µm/h to 87 µm/h on plates. While growth rates generally increased with the length of the root, growth rates still varied substantially between roots of similar length but the variance was similar in imaging chambers and on plates.
Lateral roots proliferate in imaging chambers and can be imaged using high numerical-aperture objectives.
Lateral roots co-expressing the plasma-membrane marker NPSN12-YFP14 and the microtubule marker UBQ1::mRFP-TUBULIN BETA6 (RFP-TUB6)15 were imaged by CLSM10 at 1 h intervals to document microtubule behavior during cell proliferation in imaging chambers (Figure 2D, Movie 3). In these chambers, lateral roots were highly stable in their position on the x, y and z axes, allowing the acquisition of continuous time-lapse confocal stacks over several hours. Meristematic cells continuously proliferated, as is evident from the formation of preprophase bands (Figure 2D, green arrows), mitotic spindles (Figure 2D, white arrows), and phragmoplasts (Figure 2D, blue arrows). As with seedlings on Petri plates, fully elongated root cells in imaging chambers initiated root hairs (Figure 2A, Figure 2D, arrow heads), indicating further that development proceeded as expected in the imaging chambers. To provide a wide field-of-view, the images in Figure 2D were obtained with a 20x/0.7 NA objective. It was also possible to obtain high resolution images of lateral roots using a high numerical-aperture 63x/1.2 NA water-immersion objective (Figure 2E).
Lateral root growth is sustained in imaging chambers for up to three days.
To explore how long imaging chambers could support lateral root growth at physiological rates, lateral root length in imaging chambers (n = 27) and on plates (n = 33) was quantified at 0 h, 24 h, 48 h, and 72 h. The roots chosen were shorter than 200 µm at 0 h, with an average lateral root length of 117 µm in imaging chambers and 121 µm on plates. Since shorter roots grow more slowly on average (Figure 2C), they grew over the edge of the agar slab less often, and were easier to image. The average growth of all lateral roots was not significantly different in imaging chambers compared to Petri plates in the first 48 h (Figure 3A). However, the average growth was significantly reduced between 48 h and 72 h (Figure 3A). While growth rates appear to generally slow down in imaging chambers after 48 h, growth rates between individuals remain highly variable (Figure 3B, 3C), which means that there is a substantial subset of lateral roots that grows at comparable rates on plates and in chambers over the complete 72 h growth period. 23 of the 33 lateral roots grown on plates (69.7%) reach a length within one standard deviation of the mean length at 72 h (between 927 and 3,163 µm, Figure 3B, red). 10 of 27 (37.0%) lateral roots in imaging chambers reach a length within this interval (Figure 3B, red).
Imaging chambers can easily be adapted for older lateral roots, primary roots and hypocotyls.
One of the limitations of the original imaging chamber design was that while the stiff agar slab provided some mechanical resistance, gravitropically growing roots eventually penetrated the agar slab, resulting in loss of image quality (Figure 4B) and growth beyond the working distance of high NA lenses, even the relatively long working-distance (0.3 mm) 63X/1.3 NA CS2 objective. Young lateral roots lack the statolith-containing columella cells required for gravitropism17 so they initially grew parallel to the coverslip in imaging chambers (Figure 2E). As they grow longer and columella and statoliths form, lateral roots exhibit increasing positive gravitropism, while primary roots exhibit a strong gravitropic response from germination onwards. This restricts to a few hours the period over which primary roots can be imaged and it severely limits the starting-length of lateral roots that can be imaged continuously for 48 h or more. To overcome this limitation, the chamber was modified such that growth is maintained parallel to the coverslip by a cellulosic cellophane membrane that acts as a penetration barrier on the surface of the agar slab (Figure 4A). Initial attempts using a 1.5% agar slab caused reduced root growth within the first 24 h, owing perhaps to mechanical stress. Lower agar concentrations in the slab were tested and it was found that in chambers with a slab containing 0.8% agar and cellulose film, young lateral root growth rates did not differ significantly from growth rates in conventional chambers for the first 48 h. For 0-24 h, mean root growth in cellulose chambers and conventional chambers was 242 µm and 262 µm respectively (p = 0.78 Student's t-Test) and for 24-48 h was 330 µm and 355 µm1 respectively (p = 0.67 Student's t-Test); n = 12 and n = 11, respectively. This arrangement effectively prevented lateral roots from growing away from the coverslip, into the agar, and also allowed imaging of primary roots for up to 48 h (Figure 4C).
To test whether the imaging chambers could be adapted for organs other than roots, Arabidopsis hypocotyls were chosen. Hypocotyls are substantially thicker than roots so in the conventional imaging chamber, the uppermost cells were squeezed against the coverslip, leading to deformation. An alternative design was therefore developed using cavity slides that contain an oval depression in their center (Figure 4D). In this configuration, a 1 mm thick slab of agar (prepared as in 2.2 above) was positioned with one end protruding by a few mm into the slide cavity (Figure 4D). Hypocotyls were positioned above the cavity in the slide while the root was positioned above the horizontal part. This ensured that the seedling was fixed in space by the root, but the hypocotyl was not squashed. While growth rates in chambers compared to plates were not systematically quantified, hypocotyls in imaging chambers exhibited the well-described wave of longitudinal growth migrating up the hypocotyl (Figure 4E, 4F)16.

Figure 1: Assembling an imaging chamber. All details are described in the text. Please click here to view a larger version of this figure.
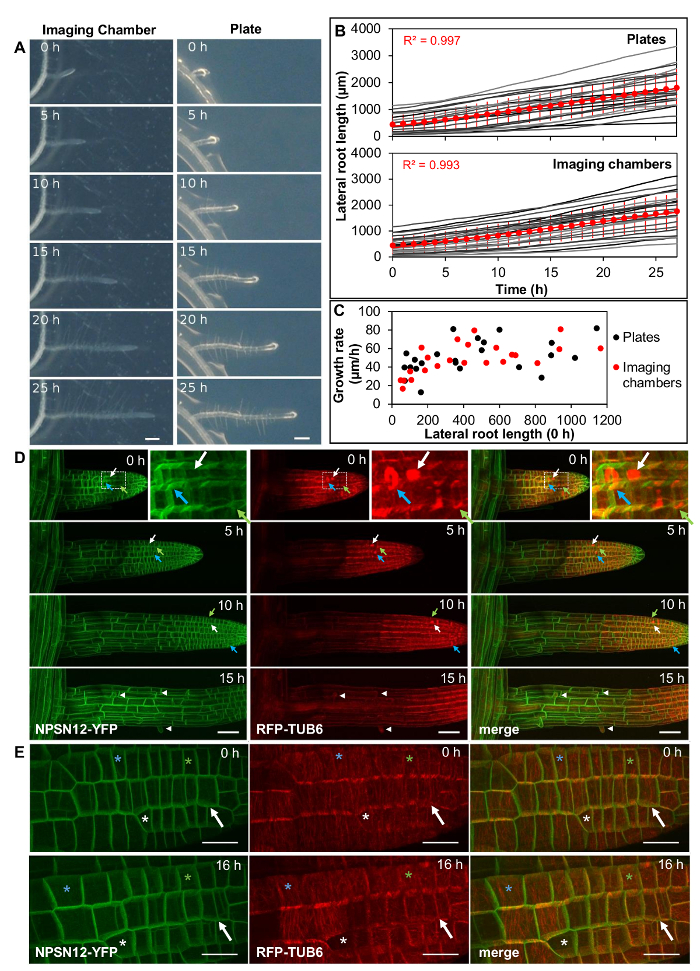
Figure 2: Lateral roots grow under physiological conditions in imaging chambers. (A) Arabidopsis lateral root development in an imaging chamber (left) and on a plate (right) over 25 h (constant light, horizontal incubation, simultaneous imaging). Scale bars = 200 µm. (B) Plot showing lateral root lengths over 27 h (measured in 1 h intervals), of lateral roots as shown in (A). Individual roots plotted in grey (n = 23 for plates and imaging chambers), average length as red circles. Error bars = 1 SD. Red line is linear fit through average lengths. (C) Plot showing average growth rate for all lateral roots shown in (B) relative to their initial length. (D) Maximum intensity projections of 3D confocal stacks acquired from lateral roots expressing NPSN12-YFP and RFP-TUB6 at consecutive time points. Inset: Magnification of boxed region. Green arrows: preprophase bands; white arrows: mitotic spindles; blue arrows: phragmoplasts; arrowheads: root hairs. (E) Maximum intensity projections of high-resolution images acquired from a young lateral root of the transgenic line shown in (D) using a 63X/1.2 NA objective and Nyquist sampling (pixel dimensions 77 nm x 77 nm) at 0 h and 16 h; asterisks locate identical cells at each time point; arrow indicates a cell that divides between 0h and 16 h. Scale bars = 50 µm (A-C); 20 µm (E). Please click here to view a larger version of this figure.
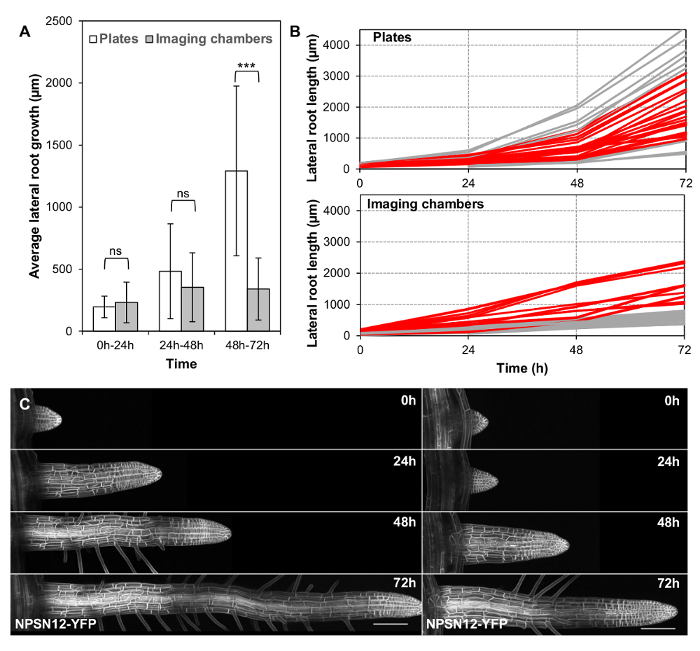
Figure 3: Long-term lateral root development in imaging chambers. (A) Plot showing average absolute root growth in three consecutive 24 h intervals in imaging chambers compared to plates. n.s. indicates p >0.05; *** indicates p <0.001 (student t-test). n = 33 (plates) and n = 27 (imaging chambers). (B) Plot showing lateral root lengths at consecutive time points on all roots used in (A). Red: all lateral roots with a final length within 1 SD of the mean of all lateral roots grown on plates (between 927 and 3163 µm). (C) Maximum intensity projections of representative 3D confocal stacks of the plasma-membrane marker NPSN12-YFP in lateral roots, acquired at consecutive time points over 72 h. Scale bars = 100 µm. Please click here to view a larger version of this figure.

Figure 4: Imaging chambers can be adapted for other plant organs. (A–C) Imaging chamber adaption for primary roots. (A) Schematic of imaging chamber design. This is identical to the standard design (Figure 1) apart from two modifications: the MS medium slab contains 0.8% agar instead of 1.5% agar, and a cellulose film (red) is placed between agar and plant to prevent growth into the agar. (B) XY (top) and XZ (bottom) maximum intensity projections of representative 3D confocal stacks of the plasma-membrane marker NPSN12-YFP in the primary root of a 7 day old seedling imaged at 0 h and 48 h in a conventional imaging chamber. Note that the root grows into the agar due to its gravitropic response. (C) XY (top) and XZ (bottom) maximum intensity projections of representative 3D confocal stacks of the plasma-membrane marker NPSN12-YFP in the primary root of a seven-day old seedling imaged at 0 h and 48 h in the imaging chamber with cellulose film. Before application, the cellulose film was sterilized in 80% ethanol and soaked in liquid ½ MS medium. Note that gravitropic growth of the root into the agar is prevented. (D–F) Imaging chamber adaptation for hypocotyls. (D) Schematic showing hypocotyl imaging chamber design. A poly(dimethylsiloxane) gum gasket (grey) is manufactured on a cavity slide between two glass strips (yellow). A slab of 1.5% agar of even thickness (beige) is placed on the slide, partially reaching into the cavity. The chamber is filled with PFD (blue). Seedlings are placed onto the agar slab so that the hypocotyl occupies the downwards-sloping region of the agar slab in the cavity while the root is positioned the horizontal part of the agar. The chamber is closed with a coverslip. (E) Maximum intensity projections of representative 3D confocal stacks of the plasma-membrane marker NPSN12-YFP in a hypocotyl of a two-day old seedling imaged over 48 h at consecutive time points. (F) Longitudinal growth in two consecutive time intervals of individual cells along the basal to apical axis (relative positions at 0 h along the axis shown in E) of the hypocotyl shown in (E). Note the previously described wave of growth migrating up the hypocotyl16. Scale bars = 100 µm. Please click here to view a larger version of this figure.

Movie 1: Please click here to download this movie.