The method described in this paper can be used to observe embryogenesis directly under a microscope, annotate the cell identity specification during embryogenesis with specific markers, and characterize the role of a particular gene during embryogenesis.
Representative results from the analysis of embryo patterning (from the elongated zygote stage to the mature walking-stick stage) in the wild-type Arabidopsis are shown in Figure 2. After ovule clearance, the embryos at different developmental stages could be distinguished by DIC microscopy (Figure 2). The expression pattern of DR5-GFP was recorded during embryogenesis under confocal laser scanning microscopy (Figure 3) to determine the distribution of auxin in the embryo and examine the potential role of auxin during embryogenesis in Arabidopsis.
This protocol can also be used to characterize the roles of specific genes during embryogenesis. Here, the role of Naa10 during embryogenesis in Arabidopsis was examined as an example. Abnormal division of the hypophysis was observed (askew and vertical division of the hypophysis in Naa10 compared with transverse asymmetric division of the hypophysis in wild-type plants, Figure 4). The distribution of auxin at the globular stage was nearly uniform in most Naa10 embryos and retained a broader signal in the hypophysis, as compared to the wild type embryos (Figure 5). These results indicate that Naa10 is required for embryogenesis, and that it may be involved in embryogenesis via the auxin signaling pathway in Arabidopsis.

Figure 1: Schematic Diagram for the Dissection of Arabidopsis Silique. (A) The silique attached on the glass slide was pasted with a piece of double-sided adhesive tape. (B) The enlarged image of the box in A. The two imaginary lines represented the location to be split. (C) The enlarged image of the split silique in the box in D. (D) The image of the split silique. Please click here to view a larger version of this figure.
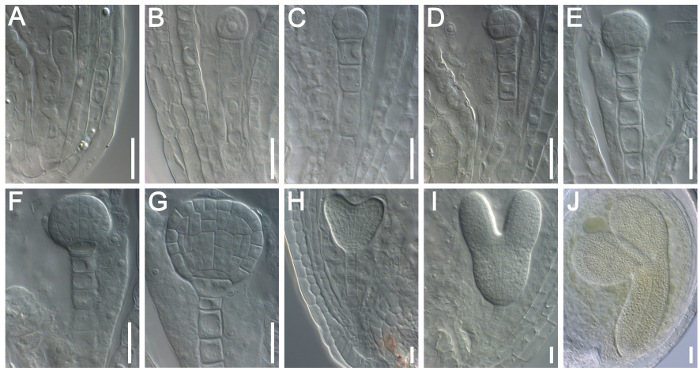
Figure 2: Examination of Embryogenesis in Wild-type (Col-0) Arabidopsis by DIC Microscopy. (A) Elongated zygote. (B) A 1-cell stage embryo at 1 DAP. (C) A 2/4-cell stage embryo at 2 DAP. (D) An octant stage embryo at 3 DAP. (E) A dermatogen stage embryo at 3 DAP. (F) An early globular stage embryo at 4 DAP. (G) A late globular stage embryo at 5 DAP. (H) A heart stage embryo at 6 DAP. (I) A torpedo stage embryo at 7 DAP. (J) A walking-stick stage embryo at 8 DAP. Scale bars = 10 µm. Please click here to view a larger version of this figure.
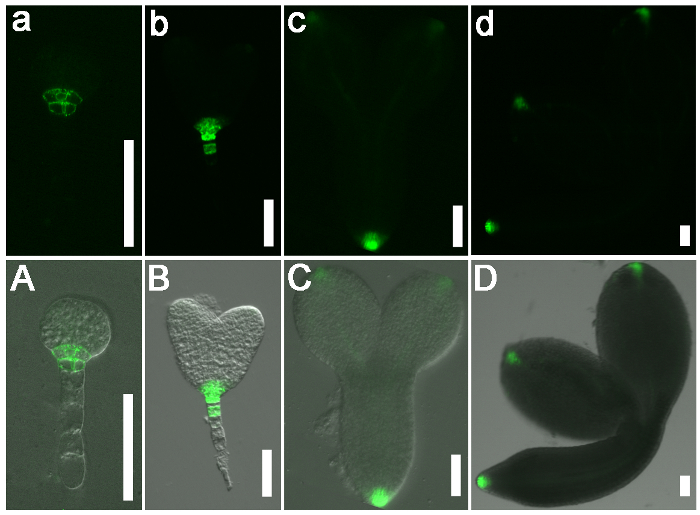
Figure 3: The Expression Pattern of DR5 in Wild-type (Col-0) Arabidopsis Embryos. (a–d) GFP signals. (A-D) Merged images with bright field images. DR5 was expressed in the root pole of the wild-type globular stage (a and A, 4 DAP) and heart stage (b and B, 6 DAP) embryos. DR5 was also expressed in the embryonic cotyledon tips (c and C, 7 DAP) and in the vasculature of mature wild-type embryos (d and D, 8 DAP). Scale bars = 50 µm. Please click here to view a larger version of this figure.
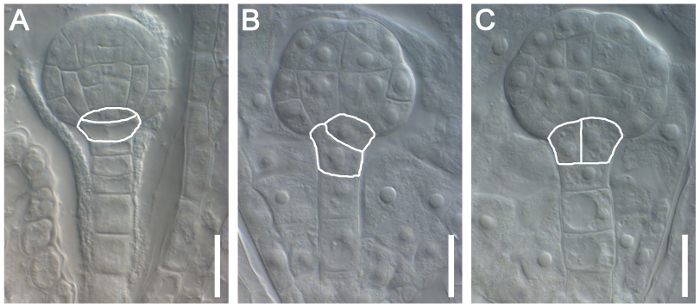
Figure 4: Characterization of the Role of Naa10 during Embryogenesis in Arabidopsis. Normal division of the hypophysis in a wild-type plant (A). Abnormal division of the hypophysis in Naa10 mutant plants is marked with a white line in (B) (askew division of the hypophysis) and (C) (vertical division of the hypophysis). Scale bars = 10 µm. Please click here to view a larger version of this figure.
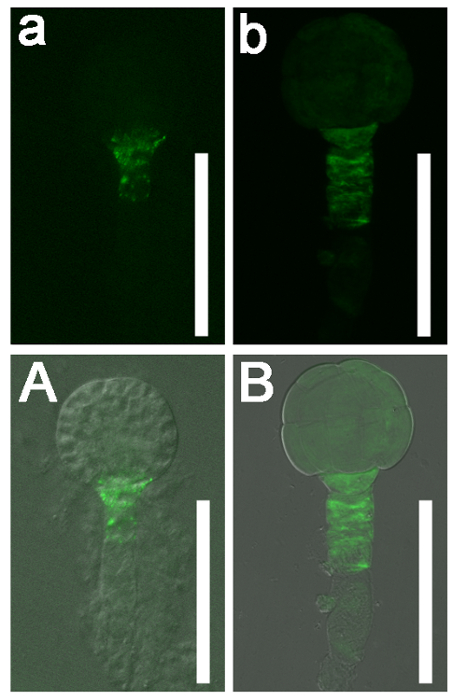
Figure 5: The DR5 Expression Pattern in Globular Stage wild-type (Col-0) Arabidopsis and Naa10 Embryos. (a and b) GFP signals. (A and B) Merged images with bright field images. DR5 was expressed in the root pole of a wild-type globular stage embryo (a and A). DR5 was nearly uniformly expressed in a globular stage Naa10 embryo and retained the broader signal in hypophysis as compared to the wild type (b and B). Scale bars = 50 µm. Please click here to view a larger version of this figure.
