1. Select a Tree for Instrumentation
- Select trees for measurement. Ideally, select trees that are healthy with a generally round stem cross-section, and a diameter between 1-2 times the tine length, or a sapwood depth greater than the length of the sensor tines (~5 cm for the specific capacitance sensors demonstrated here). Measure the depth of the sapwood using tree cores, or for many species, calculate sapwood depth through allometric equations relating sapwood area to stem diameter 29,54, as measured by a standard diameter tape.
NOTE: However, some types of capacitance sensors may be able to be trimmed to an appropriate length following Step 1.2 without adversely affecting measurement accuracy. Only capacitance sensors with rigid measurement tines that do not contain wiring can be trimmed or cut. Separate calibration is necessary for trimmed sensors. Therefore, select the tree and determine the appropriate tine length prior to the calibration procedure. - Determine the appropriate length of the sensor tines based on the tree's diameter and sapwood depth.
NOTE: Capacitance sensors integrate moisture information over the length of the sensor tines, L. It is therefore assumed that the integrated observation they provide is representative of the entire stem when the diameter is between L and 2L. The bark and phloem will not affect the measurement as they are removed from the measurement area prior to installation (see section 4.2).- Trim the sensors for trees where L is greater than the stem diameter, (see Step 1.3) such that the tines do not penetrate through the opposite side of the stem.
- Measure the combined signal from the sapwood and heartwood with the sensor for trees where the diameter is larger than 2L, but sapwood depth is less than L.
NOTE: As water content differs in these two tissue types, this may cause a bias if it is assumed that the observation is representative of the whole stem. In such cases, or when the user is only interested in the sapwood water fluctuations, the sensor tines must be trimmed to the sapwood depth such that the observations will represent the water content fluctuations of the sapwood (active xylem) only. When the sapwood depth is larger than L, observations represent only the sapwood, but the sensor does not need to be trimmed.
- If necessary (as determined in Steps 1.2.1 and 1.2.2), cut the sensor to fit for the specified application. To cut the sensor tines, clamp the sensor securely to a workbench and, while wearing proper protective equipment, use a power rotary tool equipped with a steel cutting disk to cut each tine to precisely the same length.
2. Harvest and Prepare Wood Samples of All Species of Interest to Generate a Species-Specific Calibration
- Collect a trunk, juvenile stem, or large branch at least 6 cm in diameter from the species of interest. Larger diameters are preferred in order to maximize the amount of wood encasing the sensor and most closely approximate the trunk density for field measurements. Remove all attached branches or leaves, and any lichen or loose material.
NOTE: Refer to the Discussion section for further discussion of uncertainty due to differences in branch and stem density. - Segment the stem into 25 or more cylindrical sections of ~15 cm length.
- Label each segment and record the average diameter and length of each. Approximate the segments' volume as the volume of a cylinder.
- Separate the segments into two groups for differential rehydration and drying. Place roughly 1/3 of the segments in a water bath to rehydrate, and the other 2/3 of the segments in a drying oven at 60 °C to dehydrate. Keep two segments separate for immediate measurement: preferably one from the middle of the length of the stem, and one from an end.
NOTE: Typically, segments drying at 60 °C will be fully desiccated after ~2 weeks, and fully rehydrated after ~3 days. Remove individual segments from the oven/water bath and measure daily or twice-daily intervals (see Step 3.7). In order to produce a gradient of measurements spanning the largest range of possible volumetric water contents.
3. Create a Calibration Relationship between Sensor Output and Volumetric Water Content
- Connect a capacitance sensor to a data logging device, following the instructions provided by the manufacturer, and to a computer for real-time sensor readout visualization. Set the time interval for data collection to 30 s.
- Using a premade drilling template held securely in place to maintain alignment, and a drill bit slightly smaller than the diameter of the tines of the capacitance sensor (3.57 mm for the sensor used in this experiment), drill two vertical sets of three holes with the second set located approximately 150 ° away around the wood segment with a vertical separation of a few centimeters to ensure that there is no potential overlap between holes. Use the wood segments set aside in Step 2.4 for the first set of measurements.
- Weigh the segment and record the weight to the nearest 0.01 g at the time of measurement. Cap the ends of the segments with plastic wrap to prevent additional drying.
- Immediately after weighing, clean the tines of the capacitance sensor with an alcohol swab and insert it into the stem segment completely, such that no part of the steel tines is visible. Wait for the measurement reading on the output screen to stabilize (2 or 3 min, generally). Record the sensor outputs of temperature, electrical conductivity, and dielectric permittivity (εb) every 30 s for 5 min and compute the average of the 10 measurements.
- Gently remove the sensor from the segment, clean the tines with an alcohol swab, and wait for the output readings to return to zero. Repeat the measurement procedure from step 3.3 onward in the second set of pre-drilled holes.
- Remove plastic from the ends and place the stem segment in the drying oven. Allow it to desiccate completely (generally 2 weeks, or until weight has stabilized for several days).
- Repeat steps 3.3 through 3.7 for all segments. Remove segments from the drying oven and measure more frequently (twice daily) within the first several days of drying, due to the higher rate of initial moisture loss than during the last several days (daily). Dry excess water from the surface of segments removed from the water bath using a paper towel before weighing and measurement. Measure one rehydrated stem segment per day, until all rehydrated segments have been measured.
- After complete desiccation, record the final dry weight of all stem segments.
- Calculate the volumetric water content (VWC, cm3water /cm3wood) of each stem segment at the time of measurement, using the following formula:
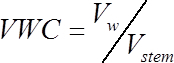 (1)
(1)
Where Vw is the volume of water (cm3), and Vstem is the volume of the stem segment (cm3) calculated in step 1.2.- Calculate the volume of water in each segment at the time of measurement as:
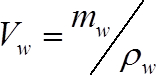 (2)
(2)
Where mw is the mass water in the stem segment (g) at the time of measurement, and ρw is the density of water (1 g cm-3) - Calculate the mass of water in each segment at the time of measurement as:
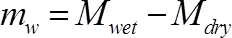 (3)
(3)
Where Mwet is the mass of the stem segment at the time of measurement (g), and Mdry is the final weight of the segment (g).
- Calculate the volume of water in each segment at the time of measurement as:
- Using a statistical analysis software package, create a best-fit linear regression between sensor-observed dielectric permittivity and VWC (Figure 1, Table 1).
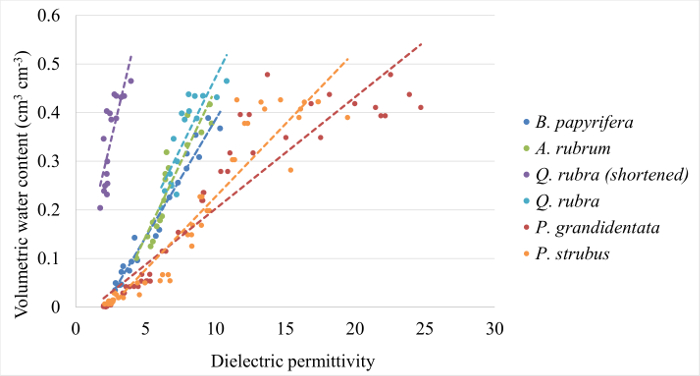
Figure 1: Example calibration curves. Calibration curves generated for Quercus rubra, Acer rubrum, Betula papyrifera, Populus grandidentata, and Pinus strobus following Parts 1 and 2 of this protocol. Equations and coefficients of determination are provided for each in Table 1. Please click here to view a larger version of this figure.
| m | b | R2 | |
| B. paprifera | 0.048 | -0.098 | 0.967 |
| A. rubrum | 0.067 | -0.158 | 0.853 |
| Q. rubra (shortened) | 0.120 | 0.041 | 0.636 |
| Q. rubra | 0.058 | -0.109 | 0.718 |
| P. grandidentata | 0.023 | -0.028 | 0.887 |
| P. strobus | 0.030 | -0.072 | 0.900 |
Table 1: Calibration equations for the conversion of εb a VWC for five temperate tree species. Coefficients 'm' and 'b' are presented for a linear equation in the standard form: VWC = m*εb+b.
4. Installing Capacitance Sensors in Trees for Field Measurements
- Prior to sensor installation, record the stem diameter and the height above the ground surface for each sensor location. Typically, to monitor VWC in the trunk, place one sensor ~0.5 m above the ground surface, and place a second one just below the first major branching split (~7.5 m above the ground, Figure 2).
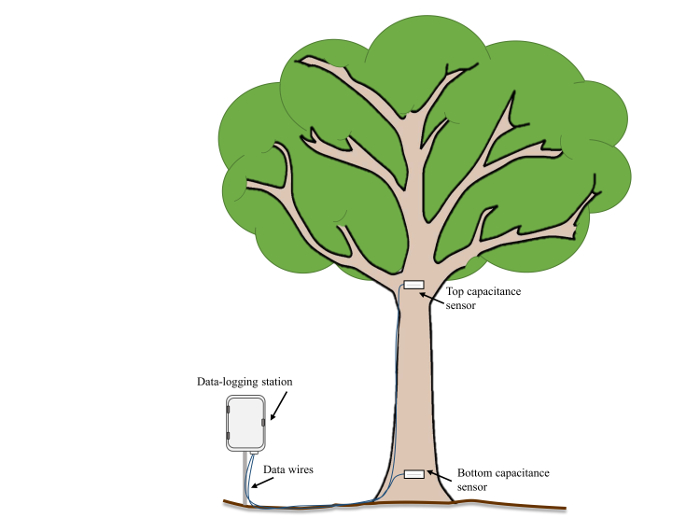
Figure 2: Example field experiment setup. A schematic diagram of sensor locations and orientation in a mature tree adjacent to a data logging station. Please click here to view a larger version of this figure.
- Remove the bark to the cambium and create a flat surface where the sensor will be installed using a draw blade. Ensure that this surface is wide and flat enough that the sensor sits flush against the surface of the tree when it is installed such that no part of the tines is exposed. Remove the bark and cambium to ensure that measurements include only water content in the xylem, while excluding water content in the bark or phloem.
- Drill the holes for the tines. Use a drill bit slightly smaller than the diameter of the tines for softer wood, while using a bit closer to the true size of the tines for harder wood (3.57 mm as used in this protocol). For high quality measurements, ensure that the sensor tines make good contact with the wood. Since there will not be a need to remove the sensor regularly, as was the case during the calibration procedure, a slightly smaller drill bit may be used here than the one used for calibration.
- Clean the sensor tines with an alcohol swab to remove any dirt or skin oils, and insert the sensor into the pre-drilled holes. If the sensor meets too much resistance upon insertion, gently back it out and re-drill the holes to widen them slightly. Ensure that all three tines are fully inserted and the body of the sensor sits flush against the tree trunk.
- Use a silicon-based sealant to seal the sensor against the tree trunk to help keep stem flow from entering the holes and to prevent pest infestation.
- Cover the sensor with reflective insulation to avoid external heating.
- Following the manufacturers' specifications, connect all sensors to a 12 V power source and a compatible data logger. Use data collection intervals of 5 min for sensors deployed in the field, but use longer intervals to conserve power in sites where power is limiting, e.g., solar/battery operated sites.
5. Process the Raw Data to Stem-Water Storage Using the Calibration Curve
- Using the equation of the calibration curve generated in step 2.9, convert the sensor output to VWC for all observations. VWC is generally expressed as cm3 /cm3 or as a percent. The percent VWC will therefore range between 0 and 100%.
- Integrate the total volume of interest (cm3) between the two measurement heights (e.g., de Figure 2: 0.5 m and 7.5 m) by assuming stem diameter changes linearly with height. For trees where the observation represents the whole stem (i.e., trees of diameters between L and 2L, using unmodified sensors, or thinner trees where the sensor was trimmed to the stem diameter, see section 2.2), use equation 4:
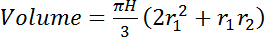 (4)
(4)
where H is the height difference (vertical distance) between the sensors (e.g., in Figure 2: 700 cm) and r1 and r2 are the radii of the stem (cm) at the bottom and top sensors' locations, respectively.
For trees where sensors provide an estimate only of the sapwood water content (see sensor length determination, section 1.2) use eq. 5:
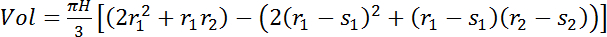 (5)
(5)
where s1 and s2 are the sapwood depths (cm) at the bottom and top sensors' locations, respectively. - Calculate the volume of stem- or sapwood- stored water by multiplying the tree's average VWC by the total volume of interest.
In this section, we present calibration data for five common eastern forest tree species, followed by a detailed analysis of field measurements of stem-water storage in three Acer rubrum individuals during the 2016 growing season. Calibration curves were generated for Acer rubrum, Betula papyrifera, Pinus strobus, Populus grandidentata, and Quercus rubra (Figure 1). Slopes of the curves differed by as much as 97.7% for P. grandidentata and A. rubrum (Table 1) demonstrating the need to perform species-specific calibration to obtain accurate VWC measurements. Volumetric water content was monitored at the base of the tree stem 0.5 m above the ground surface, and at the base of the live crown, 7.5 m above the ground surface (Figure 2). Diurnal depletion and replenishment of VWC were observed in both locations, with VWC at the top measurement point showing larger diurnal variability than the lower location (Figure 3). VWC at the top measurement location ranged from 0.55 to 0.88 cm3/cm3, while VWC at the lower measurement location ranged from 0.72 to 0.95 cm3/cm3. During transpiration, water is lost from leaves and depleted most quickly from distal branches. Therefore, it follows that VWC measured at the base of the live crown should be lower than at the base of the trunk when storage is being actively withdrawn. Measurements at both locations showed the same general trends during the growing season (Figure 3).
To evaluate the observed diurnal dynamics of changing stem water content, we calculated the time rate of change in stem-average total water content, ΔStorage (g/s), (Figure 4). ΔStorage was compared to sap flux data collected simultaneously in the same tree using Granier-style thermal dissipation sensors55. Negative values in ΔStorage indicate the rate of reduction in trunk water storage. Changes in storage began to occur shortly after dawn (6:00 am), while sap flux lagged behind between an hour and a half and two hours (~8:00 am). In general, lag-times between sap flux and transpiration are estimated at slightly more than an hour9. The slowing depletion of storage between noon and 4:00 pm can be attributed to mid-day stomatal closure, which is also responsible for the asymmetric shape of the sap flux curve throughout the afternoon56,57. The congruence in results obtained using well-established sap flux techniques and our storage measurement procedure demonstrate the ability of capacitance sensors to capture fast dynamics of changes in stem water storage in mature trees.
Three different mature Acer rubrum (diameters at breast height, DBH: 29.1 cm, 28.3 cm, 22.7 cm) in a forest research site in northern Michigan were monitored during the 2016 growing season. While total stem water storage is dependent on the volume of the trunk (Figure 5A), all three individuals exhibited patterns of recharge and replenishment consistent with seasonal trends in sap flux (Figure 5B) and available soil moisture within the top 1 m depth (Figure 5C). Stem-stored water, sap flux, and soil moisture were all at their lowest during the end-of-summer extended interstorm period, day-of-year (DOY) 200-225. At this time, sap flux and stem water content were reduced significantly in all monitored individuals. We estimated the drought recovery time in response to the observed decline in soil water content from 10% to 5.6% to be approximately 10 days for A. rubrum, based on the time required for stem-water storage to rebound to pre-drought levels.
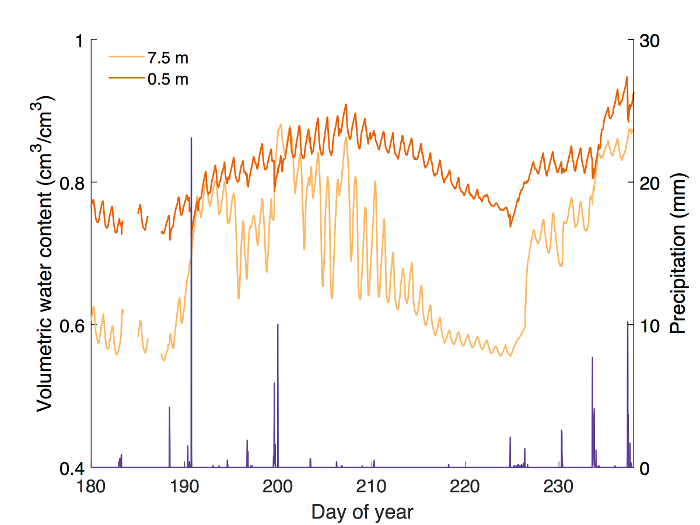
Figure 3: Example results from a field experiment monitoring stem water content at 0.5 and 7.5 m heights in Acer rubrum (DBH = 29.1 cm). Water content at 7.5 m above the ground generally remained lower than water content at 0.5 m. Diurnal fluctuations in water content at 7.5m were larger than those recorded closer to ground level. Increases in stem water content at both heights corresponded to days following precipitation (shown in purple), with a large decrease in stem water content between DOY 210 and 225 of 2016 when little to no precipitation occurred. Missing data are the result of temporary power interruption. Please click here to view a larger version of this figure.
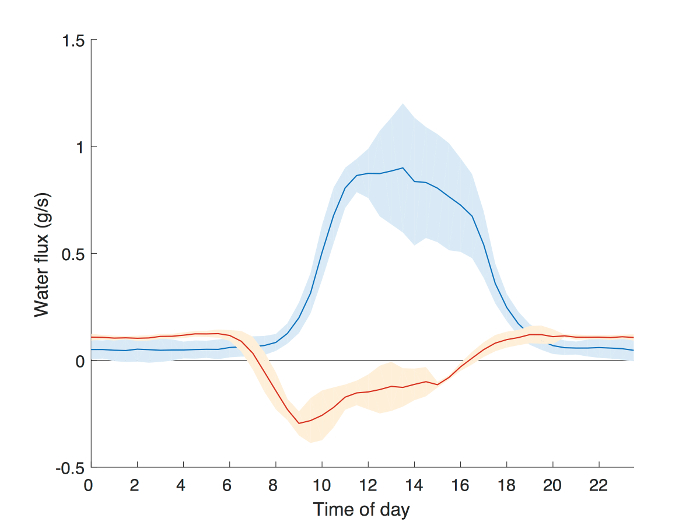
Figure 4: Example results from a field experiment comparing the time lag in water flux as measured by sap flow and the rate of change of stem water content. Sap flow was monitored simultaneously with stem water content in an Acer rubrum following the methodology outlined in Matheny, et al.29 The average diurnal sap flow rate (blue) and the time derivative of stem water storage (ΔStorage, orange) as calculated following Matheny, et al.5 are shown for 5 days when soil moisture was non-limiting (DOY 245-250, 2016). Shaded areas represent the standard deviation across the 5-day window. ΔStorage begins to decline shortly after dawn (6:00), while sap flux begins to increase roughly 2 h later (8:00). Please click here to view a larger version of this figure.
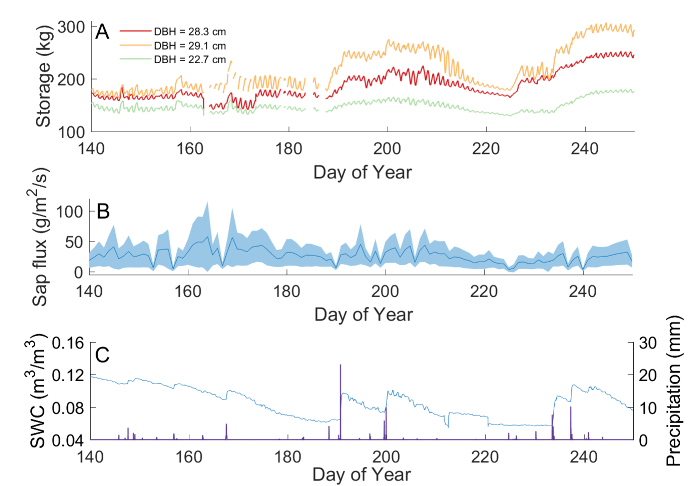
Figure 5: Example results from a field experiment monitoring stem water storage in three Acer rubrum over the course of 110 days. Stem water storage was monitored in the trunks of three Acer rubrum of different trunk sizes throughout the course of the 2016 growing season (A). Seasonal trends in stem water storage corresponded to trends seen in sap flux monitored in 15 Acer rubrum in the same forest following the procedures of Matheny, et al.29 (B). Shaded area corresponds to the standard deviation between individual trees, while the solid line represents the mean daily integrated sap flux. Both stem water storage and sap flux from Acer rubrum followed the patterns of soil water content (SWC), as integrated over the top 1 m of soil depth, and precipitation (C). Missing data are the result of temporary power interruption. Please click here to view a larger version of this figure.
