For the selection system, a sharp decrease in OD600 for strains harboring the rare-codon-rich antibiotic resistance gene should be observed in comparison to the strain harboring the wild-type antibiotic resistance gene when cultured in a suitable medium (Figure 1a). Under the same conditions, the decrease in cell OD600 becomes more obvious as the number of rare codons in the antibiotic resistance gene increases (Figure 1a). It should be noted that the inhibition of rare codon on protein expressions mostly takes place under starved conditions. Therefore, if the LB medium is not properly diluted, no significant decrease in cell OD600 will be observed for the strain harboring the rare-codon-rich marker gene in comparison to the strain harboring the wild-type gene (Figure 1b). After extra feeding of the corresponding amino acid, the OD600 for the strain harboring the rare-codon-rich antibiotic resistance gene will increase significantly and approach that of the strain harboring the wild-type gene (Figure 1c).
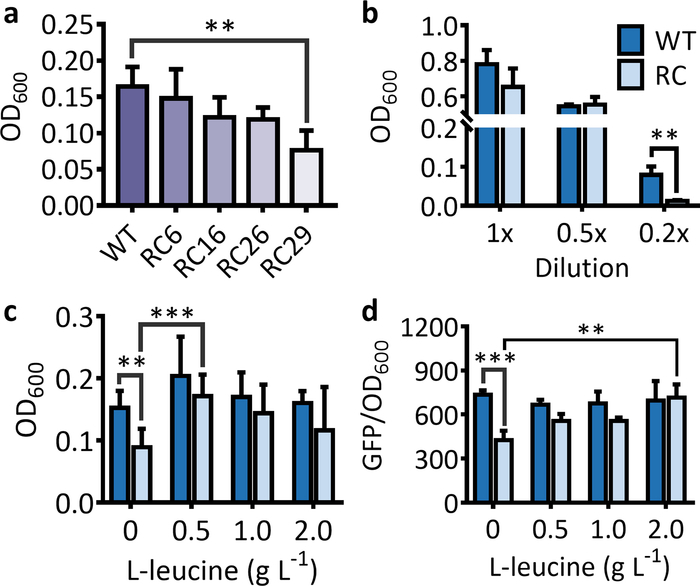
Figure 1: Effects of rare codon on the expressions of marker genes used for the selection and the screening systems. (a) The cell OD600 for an E. coli strains harboring the antibiotic resistance gene (kanR) with 6, 16, 26, and 29 leucine rare-codon (RC6, RC16, RC26, and RC29) replacement after 5 h of incubation. (b) The cell OD600 for an E. coli strain harboring the wild-type (WT) and the rare-codon-rich kanR (RC) in 1x, 0.5x, and 0.2x LB media after 5 h of incubation. (c) Effects of feeding L-leucine on the cell growth for E. coli strains harboring the leucine rare-codon-rich kanR gene after 5 h of incubation. The values and error bars represent the mean and the SD (n = 6). The feeding of L-leucine significantly increased the OD600 for cells harboring the rare-codon-rich kanR. The only exception was for the feeding of 2 g·L-1 L-leucine due to a high SD in OD600 for the feeding treatment. (d) Effects of rare-codon and L-leucine feeding on GFP expressions from the wild-type (WT) and the leucine rare-codon-rich (RC) genes after 16 h of incubation. The feeding of 0.5–2 L-1 L-leucine significantly increased the fluorescence intensity for cells harboring the rare-codon-rich gfp. The values and error bars represent the mean and the SD (n = 3). **P < 0.01, ***P < 0.001 as determined by two-tailed t-test, and only the most significant results were shown. Please click here to view a larger version of this figure.
For the screening system, the fluorescence intensity and the number of fluorescent cells will be significantly lower for the strain that expresses the fluorescent protein from the rare-codon-rich gene than from the wild-type gene (Figure 1d and Figure 2). When using the purple protein, the color developed from the rare-codon-rich ppg should be lighter than that from the wild-type gene when expressed under the same conditions for the same incubation period (Figure 3). Feeding of the corresponding amino acid will restore protein expressions from the rare-codon-rich genes. For strains harboring the rare-codon-rich gfp, the fluorescence intensity (Figure 1d) and the number of fluorescent cells (Figure 2) should increase significantly and approach that of the strains containing the wild-type gfp. When undiluted LB is used, the amino acids in the medium would be sufficient to allow slow expression of the rare-codon-rich ppg even without extra L-leucine feeding, and the expressed purple protein would become visible once the cells are pelleted (Figure 3). However, this does not conceal the fact that gene expression from the rare-codon-rich ppg was dramatically enhanced by feeding of the L-leucine to 2 g·L-1, especially when observed in liquid culture (Figure 3). Therefore, the liquid culture is a better choice for screening based on chromogenic proteins, and the use of diluted LB medium would bring a more significant difference between the phenotypes induced by the wild-type and the rare-codon-rich genes.
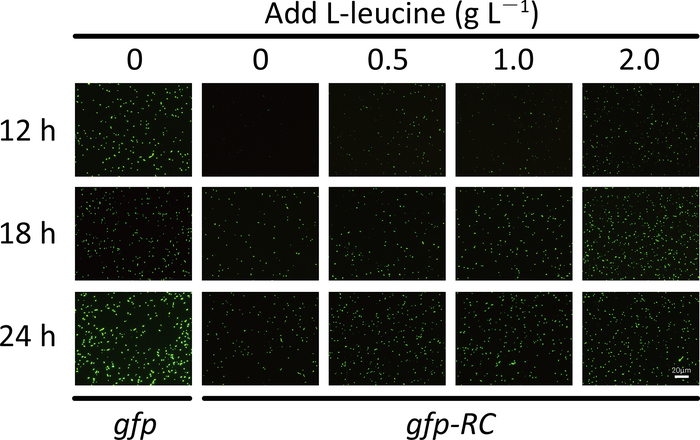
Figure 2: The number of fluorescent E. coli cells that harbor the wild-type gfp or the leucine rare codon-rich gfp (gfp-RC) after the addition of L-leucine. Cells were cultured in 1x LB medium. Scale bar = 20 μm. Please click here to view a larger version of this figure.
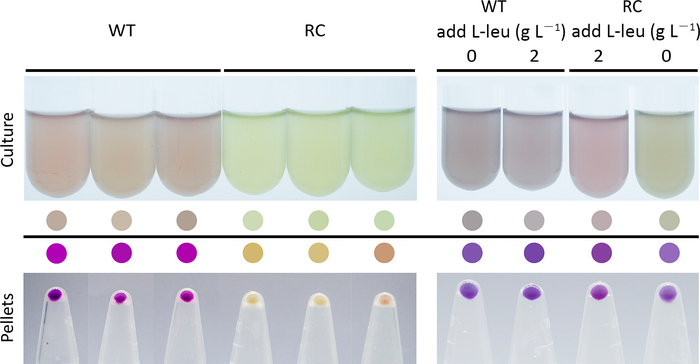
Figure 3: Color development for cells harboring the wild-type (WT) and the leucine rare-codon-rich (RC) ppg genes that encode a purple protein in 1x LB medium (left panel) and the effect of L-leucine feeding on cell culture color development (right panel). The ppg genes were induced when the cells entered the exponential phase and the images were captured 3 h after the induction. The L-leucine was added to the medium together with the inducer in the feeding assay. The colored circles were generated by picking the colors of the cell cultures and the cell pellets. Please click here to view a larger version of this figure.
The rare-codon-based strategy is able to identify overproducers of the targeted amino acids from the mutation library, and these mutants should produce higher amounts of the targeted amino acids than the parent strains (Figure 4).
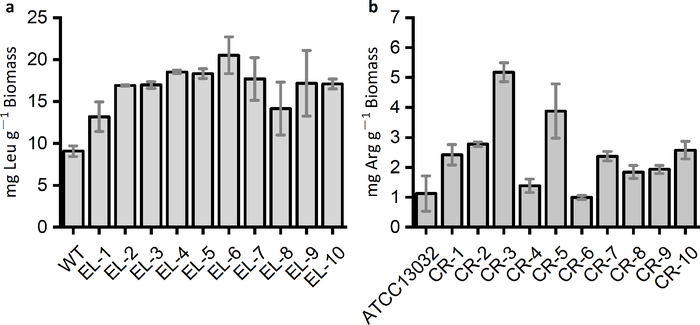
Figure 4: Amino acids produced by the wild-type and the mutated strains identified by the rare-codon-based strategy. (a) L-leucine productions of E. coli strains identified from mutation libraries by the kanR-RC29 (EL-1 to EL-5) and the gfp-RC that harbors 29 and 19 leucine rare codons (EL-6 to EL-10), respectively. (b) L-arginine productions of Corynebacterium glutamicum strains selected by the rare-codon-rich kanR, which contained eight arginine rare codons (AGG). The marker gene was introduced into the C. glutamicum mutation libraries derived from the wild-type strain ATCC13032. The selection medium was 0.3x CGIII supplied with 25 μg·mL-1 kanamycin. Please click here to view a larger version of this figure.
