NOTE: All steps in this protocol are performed at room temperature (~20-22 °C) or in constant temperature incubators set to 16 °C or 20 °C. Male and hermaphrodite C. elegans are grown using standard culture conditions and NA22 or OP50 E. coli as a food source21,22. Wild-type N2 hermaphrodites and fog-2(q71) males are used in the procedure below.
1. Day 1: Picking L4 Stage Hermaphrodites for Mating
- To obtain consistent results, all hermaphrodites should be synchronized as actively reproducing adults. Pick 20-30 L4 stage hermaphrodites to a 6 cm seeded nematode growth medium (NGM) plate. Incubate the hermaphrodites at 20 °C for 28-30 h.
NOTE: Only 12-15 hermaphrodites will be used for mating. The remaining hermaphrodites are surplus.
2. Day 1: Staining of Males with Fluorescent Mitochondrial Dye (Mito-dye)
- Make a male staining plate by placing a dot of E. coli (food dot) at the center of an unseeded NGM plate. To make the food dot, use the end of a glass stirring rod to scrape E. coli from the bacteria lawn of a seeded plate and deposit it on the unseeded plate. The dot should be ~ 5-7 mm in diameter.
- Mix together 2 µL of 1 mM mito-dye (see Table of Materials) in DMSO and 10 μL of M9 buffer (3 g of KH2PO4, 6 g of Na2HPO4, 5 g of NaCl, 1 mL of 1 M MgSO4, H2O to 1 L. Add MgSO4 after autoclaving). Pipette all of the mito-dye solution onto the food dot on the male staining plate. Let the plate dry in the dark (~30 min).
NOTE: Mito-dye is light sensitive. Shield all solution, plates, and worms containing mito-dye from light. Store the 1 mM stock at -20 °C. - Pick ~100 1-3 day old adult males23 to the mito-dye stained food dot on the male staining plate. Wrap the plate in aluminum foil and incubate overnight at 16 °C. For mating, use ~50-60 males per 12-15 hermaphrodites. If more than ~100 males are needed, make more staining plates to prevent overcrowding of males.
NOTE: Males can also be stained by incubating in a 10 μM mito-dye solution in M9 buffer for 3 h on a watch glass. Keep the worms covered to prevent evaporation and light exposure. After 3 h, use a Pasteur pipet to transfer males onto a 10 cm seeded NGM plate. Transfer as little of the mito-dye solution as possible. Wrap the plate in aluminum foil and incubate overnight in 16 °C.
3. Day 2: Mating
- Pick the stained males from Day 1 onto a new, seeded NGM plate. Leave the plate in the dark until mating. This step ensures that excess mito-dye stained bacteria around the males are removed. Carryover of excessive mito-dye stained bacteria onto the mating plate can stain hermaphrodite tissue.
- Make a mating plate by dropping 2 μL of a thick E. coli mixture on an unseeded NGM plate. Let the thick bacteria dry to make the mating dot. Males and hermaphrodites will be transferred onto this dot for mating. To make thick E. coli, spin down 3 mL of overnight E. coli and resuspend the bacteria pellet in 1 mL of M9. This mixture can be stored at 4 °C and reused for up to 6 months.
NOTE: The thickness of the E. coli solution may be adjusted. If the solution is too thin, males may crawl away from the mating dot instead of aggregating on it for mating. Mating dots made from an E. coli soluton that is too thick may decrease the mating efficiency. - While the mating plate from Step 3.2 is drying, mix together 300 μL of 1% (w/v) Tricaine (Tri), 300 μL of 0.1% (w/v) Tetramisole (Tet), and 900 μL of M9.
NOTE: Store 1% (w/v) Tricaine and 0.1% (w/v) Tetramisole as aliquotes at -20 °C. Avoid repeated freeze thaw. - Transfer 600 μL of the Tet/Tri solution to a watch glass.
- Transfer 12-15 hermaphrodites picked on Day 1 to the Tet/Tri solution in the watch glass. Incubate for 30 min to immobilize the hermaphrodites. Keep the watch glass covered to prevent the Tet/Tri solution from evaporating.
NOTE: It is important that hermaphrodites are anesthetized for at least 30 min. Less time could result in a moving worm during image acquisition, which can interefere with imaging. - While hermaphrodites are incubating, pick 50-60 stained males from Step 3.1 onto the mating dot (Step 3.2). Store the plate in the dark until Step 3.8.
- After the 30 min incubation in the Tet/Tri solution, use a glass Pasteur pipet to transfer the immobilized hermaphrodites from the watch glass onto an unseeded NGM plate. Remove as much liquid as possible and let the excess liquid dry.
NOTE: Do not let the hermaphrodites dry excessively. As soon as all visible liquid has evaporated, begin the next step. - Transfer the anesthetized hermaphrodites from the unseeded NGM plate onto the mating dot with the stained males. Incubate in the dark for 30 min to allow the males to mate with the hermaphrodites.
- After mating for 30 min, mount the hermaphrodites immediately for time-lapse imaging or transfer the hermaphrodites onto a new, seeded NGM plate to rest for 1 h before imaging.
NOTE: Time-lapse videos of the hermaphrodite uterus are used to quantify sperm velocity and reversal frequency. Still images of the uterus taken 1 h after mating are used to quantify sperm distribution, or spem guidance.
4. Day 2: Mounting Worms for Visualization
- Creating a mounting pad with 2% agarose in H2O
NOTE: 2% agarose can be made in bulk, aliquoted into glass test tubes, and stored at 4 °C. When needed, each aliquot can be microwaved before each use and stored in a heat block to prevent it from solidifying.- To make the mounting pad, align three glass microscope slides side by side with the long edges touching. Place two pieces of masking tape on top of each other on both of the outer slides. These outer glass slides with tape will act as the support so the thickness of the resulting agarose pad will be “two tape deep”.
- Place ~75 μL of melted 2% agarose on the center slide (this is the slide without tape). Immediately place a new glass microscope slide on top of the agarose. This top glass slide should be perpendicular to the other slides, with each end resting on the tape of the two support slides.
- Let the agarose harden (~30 s). Carefully remove the top glass slide by sliding it off the agarose pad.
- Place 10-15 μL of the Tet/Tri solution onto the 2% agarose pad. Transfer the mated hermaphrodites onto the pad. Take care to transfer as little bacteria as possible.
- Place a cover slip over the worms on the agarose pad.
5. Day 2: Image Acquisition Setup
NOTE: Any upright miscroscope equipped with epi-fluorescence, 10x and 60x objectives, and a digital camera can be used to acquire images for sperm distribution. Software capable of acquiring time-lapsed images are required for assessing sperm speed, directional velocity, and reversal frequency.
- Image acquisition 1 h after mating
- Mount the slide onto the microscope stage. Look through the eye pieces to scan for worms on the agarose pad using the 10x objective with the red fluorescence emission filter (TRITC filter). Once a worm has been found, briefly turn on the fluorescence light to see if the worm has mated. If sperm is visible within the uterus, switch to the 60x objective.
NOTE: The pressure created by the 60x objective on the coverslip may damage some fragile worms, causing the intestine or gonad to extrude from the animal. Scanning for successful mating using the 10x objective can minimize the worms’ exposure to the added pressure. Do not expose the mated worms to extended periods of fluorescent light. - Using Differential Interference Contrast Microscopy (DIC), position the worm so that both the vulva and one spermatheca are in view. Focus the image by focusing on the center of the spermatheca. Check the exposure for both DIC and TRITC channels. In DIC, internal worm structures should be clearly visible. In TRITC, individual sperm should be visible as distinct puncta.
NOTE: Each image should capture the uterus from the vulva to one of the spermatheca. If the uterus is too long to fit on one image, two separate images can be taken. It is not necessary for all of the images to be taken at the same exposure level. However, it is important that individual sperm can be distinguished and quantified in the fluorescence images. - Acquire DIC and fluorescence images for each uterus.
- Repeat Steps 5.1.1-5.1.3 until all mated hermaphrodites have been imaged.
- Mount the slide onto the microscope stage. Look through the eye pieces to scan for worms on the agarose pad using the 10x objective with the red fluorescence emission filter (TRITC filter). Once a worm has been found, briefly turn on the fluorescence light to see if the worm has mated. If sperm is visible within the uterus, switch to the 60x objective.
- Capturing time-lapse videos
- Scan the agarose pad and locate hermaphrodites that contain labeled sperm within the uterus, as described in Step 5.1.1 and 5.1.2
- Configure the software to acquire time-lapse images in DIC and TRITC channels. Generally, time-lapse images are taken at 15-30 s intervals for 10-20 min per uterus.
6. Quantification
- Quantifying sperm distribution on uterus images taken 1 h after mating
- Starting with the vulva on one end and the spermatheca on the other, divide the uterus into thirds. These will represent the three zones. Zone 1 (Z1) contains the vulva and Zone 3 (Z3) contains the spermatheca.
- Manually count the number of sperm within each third of the uterus, and report the number in each zone as a percent of the total sperm in the entire uterus. An example is provided below.
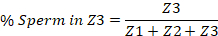
NOTE: Sometimes, the signal intensity of the TRITC channel image needs to be adjusted so that every sperm that has been captured in the image can be visible and quantified.
- Tracking sperm in time-lapse images
NOTE: In this paper, we used the NIS-Elements software for analysis. In the sections below, we give instructions for manually tracking sperm using this software (step 6.2.1) as well as the open source software ImageJ/Fiji (step 6.2.2).- Tracking sperm with NIS-Elements
- Open the .nd2 file with the time-lapse series to be tracked. To begin tracking, open up the Tracking panel by right clicking in the software and selecting Analysis Controls | Tracking.
- In the tracking panel, select Define New ROI. Define each region of interest (ROI) by clicking over each sperm that will be tracked. A colored mark will appear over the selected sperm. Click Finish when all ROIs have been selected.
- Once the ROIs have been identified, move to the next frame in the time-lapse series. Drag the ROI marker to the new position of the sperm in the image. Continue doing this until the sperm can no longer be tracked. Dotted lines will appear connecting each of the locations the ROI marker has been placed through all the frames of the time-lapse image.
NOTE: Only sperm in Zone 2 should be tracked as sperm in Zones 1 and 3 tend to move in a circular pattern even in wild-type animals. - Export all quantifiable data (e.g., path length, time, XY position, etc.) from the tracked sperm to an Excel document by clicking Export in the Tracking panel.
- Tracking sperm with Fiji
- Convert the TRITC channel images in the time-lapse series to .tif files. Save all files from one series into one folder.
- Import the images to Fiji by using the BioFormats import function. Import images from one time-lapse series as one hyperstack.
- Open TrackMate in Fiji24 via Plugins | Tracking | Manual tracking with TrackMate. A diaglogue box will open.
- Select the TrackMate tool in the Fiji toolbar. Double click on the sperm that will be tracked. A green circle with dashed lines will appear. This circle may be repositioned by clicking inside the circle and dragging to the desired position. The size of this circle may be changed by simultaneously pressing the ALT key and scrolling the mouse.
- Once the size and position of the tracker has been set, click on the circle again. The dashed green lines will turn into a solid green line. Simultaneously hit the SHIFT and L keys to turn on tracking mode. This will be indicated in the Fiji toolbar.
- Move to the next frame in the time-lapse series. To set the new location of the tracked sperm in the new frame, hover the mouse over the new point and press the A key. The tracker will now appear at the new location, and a line will appear connecting the locations where the tracker has been placed in the previous frames.
- After the traces have been completed, click Analyze in the TrackMate dialogue box to generate the data needed.
- To calculate the speed, divide the total path length of the sperm by the elapsed time.
- To calculate the vectorial velocity, draw a line through the uterus starting from the vulva pointing toward the spermatheca. Measure the distance the sperm has migrated along this line from the beginning to the end of the trace. Divide this distance by the elapsed time. Negative values indicate the sperm has migrated away from the spermatheca.
- To record reversal frequency, count the number of times during which the sperm trace has generated an angle less than 90° during three consecutive time-lapse frames.
- Tracking sperm with NIS-Elements
To generate the results depicted in this paper, fog-2(q71) males were stained with the mito-dye and mated to wild-type, N2 hermaphrodites. Figure 2 provides an overall scheme for the method, including worm preparation, mating, and analysis. As Movie 1 shows, the adult hermaphrodite reproductive tract has two arms that are mirror images of each other. Upon mating, labeled sperm are deposited in the hermaphrodite uterus through the vulva. The sperm move around the developing embryos within the uterus toward the spermatheca, where they are stored until fertilization. As germ cells in the adult hermaphrodite develop into oocytes, the proximal, most mature oocyte is pushed into the spermatheca via sheath cell contrations. Fertilization occurs while the oocyte is in the spermatheca.
To quantify sperm distribution and migration through the female reproductive tract, the hermaphrodite uterus is divided into three zones (Movie 1, Figure 3A). Zone 1 spans the first third of the uterus, starting from the vulva. Zone 2 spans the middle third of the uterus, and Zone 3 spans the last third of the uterus and includes the spermatheca. Proper sperm guidance using wild-type, N2 hermaphrodites and fog-2(q71) males should result in approximately 90% of the labeled sperm reaching the spermatheca, or Zone 3 (Figure 3B). Matings that result in too few (Figure 3C, less than 10-15 sperm) or too many (Figure 3D, uterus filled with sperm) sperm in the uterus should not be counted. In matings that result in less than 10-15 sperm, 3-4 rogue sperm may heavily skew the data. Similarly, when the uterus is filled completely with sperm, the sperm cannot migrate appropriately. Sperm may seem scattered throughout the uteri of some mutants that display poor sperm guidance phenotype. However, in this case, sperm should not fill each crevice of the uterus, as seen in Figure 3D. Quantification of each arm of the gonad is considered one sample, or one n.
Time-lapse images are taken to quantify sperm velocity and reversal frequency. Only sperm in Zone 2 should be tracked (Figure 4A) because sperm in Zones 1 and 3 (Movie 1) tend to move in a circular pattern even within wild-type animals. Time-laspe images taken at 15-30 s intervals are usually used to track sperm. Only sperm that can be followed in consecutive frames for more than 2.5-3 min are quantified. In Figure 4B-M, the sperm marked by the red and blue dots satisfy this criterion, while the sperm marked by the green dot does not. Therefore, the values defined in Figure 4N are quantified for the sperm marked by the red and blue dots (Figure 4O), while those for the sperm marked by the green dot were not quantified.
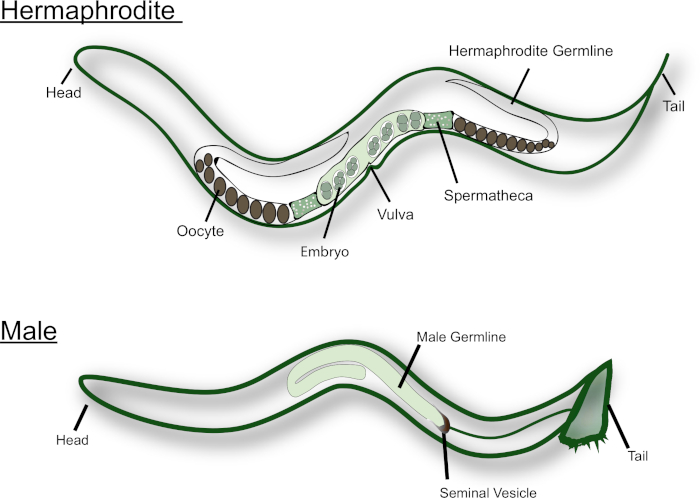
Figure 1: Cartoon of the adult C. elegans hermaphrodite and male. Major reproductive structures are labeled in the figure. Please click here to view a larger version of this figure.
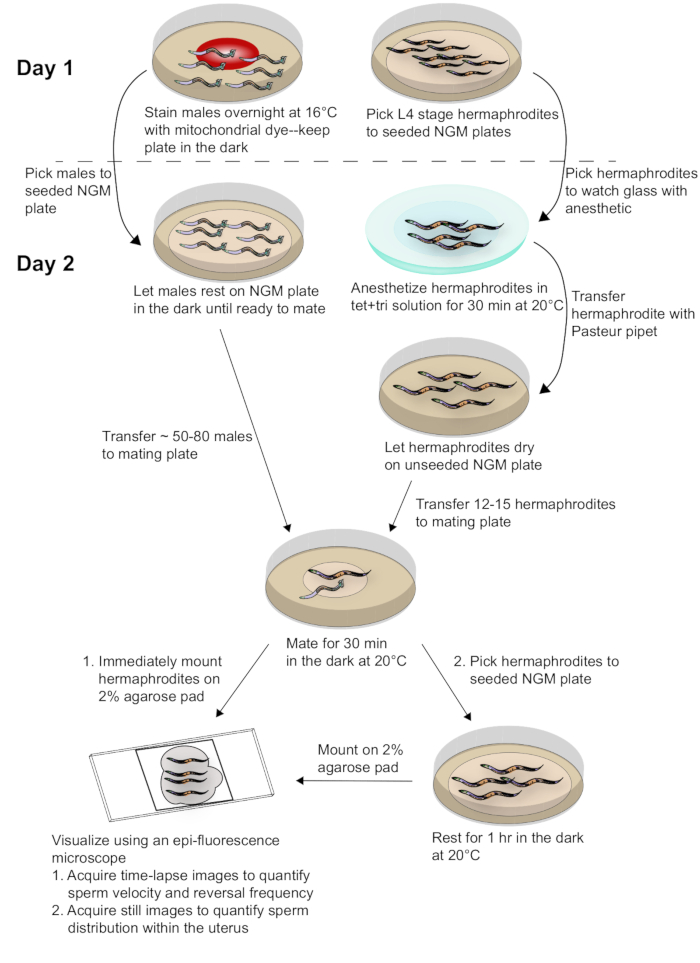
Figure 2: Schematic diagram of sample preparation and data acquisition. Males stained with the mito-dye are mated to synchronized adult hermaphrodites. Time-lapse images of mated hermaphrodites are taken immediately after mating to capture data for sperm velocity and reversal frequency. Still images of mated hermaphrodites are taken 1 h after mating to assess sperm distribution within the uterus. Refer to the text for more details. Please click here to view a larger version of this figure.
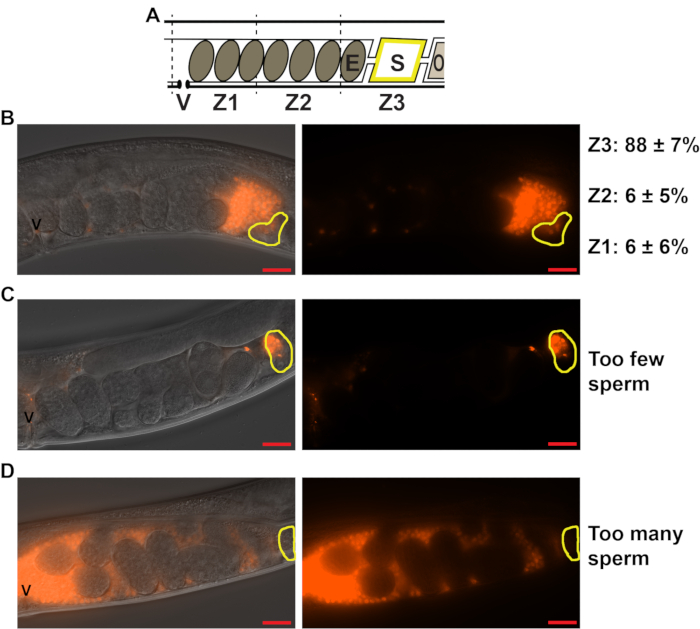
Figure 3: Quantifying sperm distribution within the hermaphrodite uterus. (A). Schematic of the C. elegans hermaphrodite uterus. V = vulva, E = embryo, S = spermatheca, O = oocyte, Z1-Z3 = Zones 1-3 used to measure sperm distribution. (B-D). DIC+TRITC merged (left panels) and TRITC only (right panels) images of the wild-type hermaphrodite uteri 1 h after mating to fog-2(q71) males stained with the mito-dye. Sperm appear red. Yellow outlines indicate the location of the spermatheca. Scale bar: 20 μm. Z1, Z2, Z3 quantification in B represent the percent sperm in each zone ± standard deviation. Images in C and D represent matings that have resulted in too few (C) or too many (D) sperm for quantification. Please click here to view a larger version of this figure.
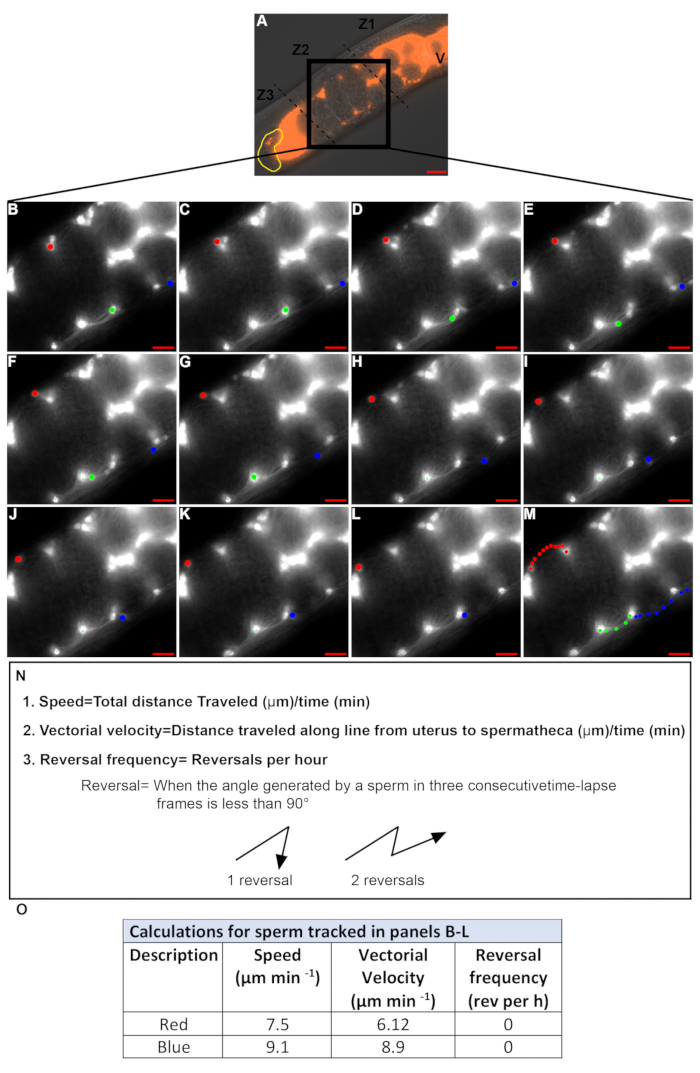
Figure 4: Quantifying sperm velocity and reversal frequency during migration through the uterus. (A). DIC+TRITC merged image of a hermaphrodite uterus containing fluorescent sperm (red). V = vulva, yellow = spermatheca, Z1-Z3 = three zones of the uterus, black box: zone 2. (B-M). Time-lapse TRITC channel images zoomed in on zone 2 (black box in A). Images were acquired at 20 s intervals. 3 individual sperm were tracked in each image (red, green, and blue dots). Colored dots in panel M represents the path of each sperm from B-L. Scale bar = 20μm. (N). Equations and definitions for sperm speed, vector velocity and reversal frequency. (O). Speed, vectorial velocity and reversal frequency of sperm tracked in panels B-L by the red and blue dots. Please click here to view a larger version of this figure.
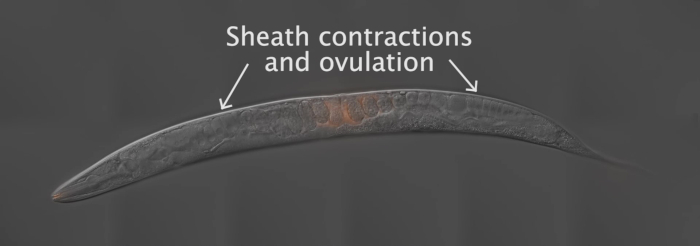
Movie 1: Movie of sperm movement and migration. A wildtype hermaphrodite was mated to fog-2(q71) males stained with mito-dye. The movie is a composite of time-lapse images taken at varied time intervals. Please click here to view this video. (Right-click to download.)
