1. Field experiment to determine the effects of UV-reflective mulch, kaolin, and companion plants on flower thrips and their minute pirate bug predator
- Establish a field experiment with a split-split-plot treatment arrangement in a randomized complete block experimental design with mulch type as whole plot treatments, kaolin and no kaolin as subplot treatments, and companion plants and no companion plants as sub-subplot treatments (Figure 1A,B)9,14.
- Layout blocks of tomatoes or pepper that are each at least 6 m wide and 72 m long.
- Randomly lay in each block whole plots of black and UV-reflective mulch, with each whole plot consisting of six raised mulch beds at least 36 m long.
- Plant one linear row of tomatoes every 45 cm or two linear rows of peppers every 30 cm into the four inner beds of each whole plot.
- Kaolin treatment
- Randomly divide each whole plot into equal subplots of kaolin or no kaolin treatments.
- Apply kaolin once or twice weekly at the rate of 7.0 kg/ha to the tomato or pepper plants in the subplots assigned to receive kaolin treatment.
- Companion plants
- Randomly divide each subplot into equal sub-subplots of companion plants or nor companion plant treatments.
- Plant two linear rows of Bidens alba (L.) every 30 cm or one linear row of Helianthus annuus L. every 30 cm into the two outer beds of each sub-subplot treatment with companion plants.

Figure 1: Example experimental field study.
(A) Randomized complete block design to evaluate the separate and interactive effects of companion plants, mulch, and kaolin effects on flower thrips and minute pirate bugs. (B) Bidens alba (L.) evaluated as a companion plant species with tomato as the crop9. Helianthus annuus L. evaluated as a companion plant species with pepper as the crop14. Please click here to view a larger version of this figure.
2. Flower thrips sampling protocol
- Prepare 50 mL sample vials before going to the experimental plots.
- Place a label with the mulch, kaolin, and companion plant treatment, the block number, and the sample date on the outside and inside of each vial.
- Put exactly 30 mL of 70% alcohol in each 50 mL vial.
- Place the vials into a tray.
- Take the trays to the experimental field site.
- Sample the flowers for thrips and minute pirate bugs.
- Randomly assign the tomato or pepper plants to be sampled in each sub-subplot.
- Sample between mid-morning and mid-afternoon.
- Take the samples from the upper half of the plant.
- Remove the vial lid. Using a sharp-edged razor or scissors, carefully remove the flower from the plant. Quickly place the flower into the appropriate pre-labeled vial. Push the flower into the alcohol of the vial (Figure 2). Replace the lid.
- Collect 10 flowers per sample. Make sure each vial is tightly sealed, then shake each vial to ensure that the flowers are within the alcohol.
- Return the trays with samples to the laboratory for storage. To ensure samples do not deteriorate prior to processing, keep samples cool and dry. Refrigerate, if possible, especially for samples that are not processed quickly.
- Repeat the sampling of each sub-subplot at least weekly during the flowering period of the crop.

Figure 2: Sample removal technique.
A sample of 10 tomato flowers being collected from a sub-sub plot in the tomato push-pull experiment9. Please click here to view a larger version of this figure.
3. Sample processing in the laboratory
- Extract the thrips and minute pirate bugs from the flowers in each sample.
- Remove the sample from the refrigerator and tray without disturbing the contents.
- Remove the lid of the vial and carefully extract with a pipette any excess alcohol above the flowers.
- Reseal the vial and shake to dislodge the thrips and minute pirate bugs in the flowers.
- Open the vial and pour the contents into a Petri dish. Rinse the inside of the vial with 70% alcohol and pour the contents into the Petri dish. Make sure all thrips and minute pirate bugs in the vial are flushed into the Petri dish.
- Dissect each flower with forceps and rinse with 70% alcohol to ensure that all the thrips and minute pirate bugs have been dislodged. Remove and discard the flower parts from the Petri dish (Figure 3).
- Transfer the Petri dish to the platform of a stereoscope with 40x–150x magnification.

Figure 3: Extracting thrips and minute pirate bugs from flowers.
A sample of 10 tomato flowers poured into a Petri dish for processing to determine the number of thrips and minute pirate bugs. Please click here to view a larger version of this figure.
- Identify and count the flower thrips in samples.
- Identify and count in each grid the number of adult males and females of each flower thrips species and the number of Frankliniella species larvae.
- Identify the adult flower thrips species in Florida based on the setae on the pronotum, head and second antennal segment10,11,12.
- Separate the adults of F. bispinosa from the adults of F. tritici and F. occidentalis by the extra stoutness of the two setae on the anterior dorsal margin of the second antennal segment (Figure 4).
- Separate the adult F. occidentalis from those of F. bispinosa and F. tritici by the near equal lengths of the anterior marginal and anterior angular major setae on the pronotum and by the longer fourth postocular setae on the head (Figure 4).
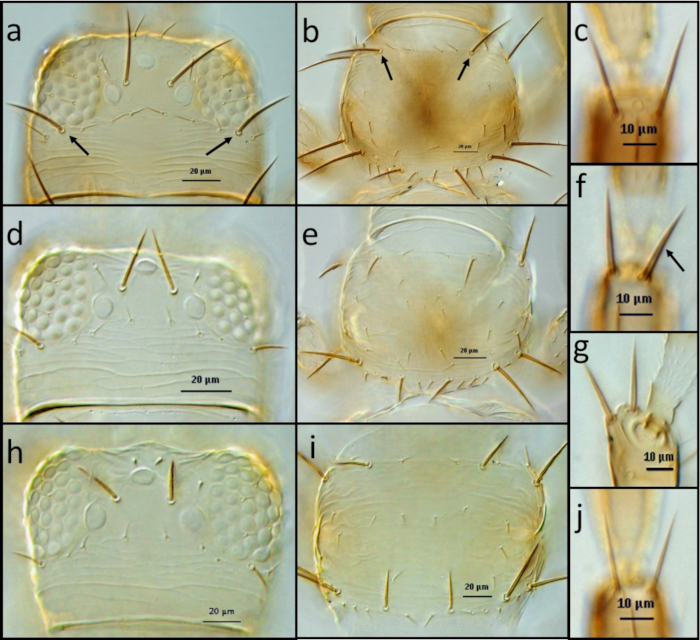
Figure 4: Examples of morphological characters to identify thrips.
(A,B,C) F occidentalis: head (A), arrows indicate postocular setae pair IV; pronotum (B), arrows indicate pair of long anteromarginal setae; distal dorsal setae of antenna segment II (B). (D,E,F,G) . F. bispinosa: head (D); pronotum (E); distal dorsal setae of antenna segment II (F,G), arrow indicates stout setae (F), lateral view of stout setae (G). (H,I,J). F. tritici: head (H); pronotum (I); distal dorsal setae of antenna segment II (J). Please click here to view a larger version of this figure.
- Identify and count the minute pirate bugs in Florida samples.
- Identify and count in each grid the number of adult O. insidiosus and O. pumilio and the number of nymphal Orius species13,15.
- Identify the adult O. insidiosus by the brown basal antennal segments, by the femora that have dark markings, and by the cuneus that is dark brown.
- Identify the adult O. pumilio by the yellow basal antennal segments, the yellow or straw-colored femora, and by the cuneus with pale straw or light-brown color.
- Add the numbers from each grid to determine the total number of adult males and females of each flower thrips species, the number of Frankliniella species larvae, the number of adult minute pirate bug of each species, and the number of minute pirate bug nymphs in the sample.
- Select representative vouchers of flower thrips and minute pirate bug adults from the samples. Label by date, plant host, location, and collector. Curate for long-term preservation.
- Transfer the data from each sample to a spreadsheet that includes the sample date, treatment, and replication.
- Create a data file that contains the data from each sample. Include the experimental location, experimental design, and the amounts and dates of each cultural practice used to establish and maintain the experiment.
- Maintain and manage the datafile with appropriate backup for long-term access.
Data collected in the study by Tyler-Julian et al.9 can be used to demonstrate the separate and combined effects of push factors (i.e., ultraviolet-reflective mulch and kaolin application) and pull factor (i.e., the companion plant Spanish needle, B. alba) on the population dynamics of F. occidentalis adult males and females in tomato flowers (Figure 1A). The agricultural plastic mulch treatments in the experiment were used to form the bed of the raised-bed plastic mulch system that is typical of the production system used to grow high-value vegetables in Florida. The mechanism of the ultraviolet-reflective mulch in pest control is a visual deterrence that disrupts host-finding by the adult thrips. Kaolin application on the tomato plants also reflects enough ultraviolet light to deter the thrips adults. Therefore, a split-split plot randomized complete block design was employed in the experiment to reduce the inter-plot interference on thrips movement resulting from the ultraviolet-reflecting properties of the mulch and kaolin treatments, with mulch treatment (ultraviolet-reflective vs. conventional black mulch) as the whole plot, kaolin treatment (twice weekly kaolin application vs. no kaolin) as the sub-plot, and companion plant treatment (companion plants vs. no companion) as the sub-subplot. Sub-subplot size was six beds by 9 m, with the four inner beds of each sub-subplot consisting of one linear row of tomato with a 45 cm spacing between plants, for a total of 80 plants per sub-subplot. Two rows of Spanish needle were planted into each of the two external beds in the sub-subplots with the companion plant with a 30 cm spacing within and between rows for a total of 128 companion plants per sub-subplot.
Two samples of 10 tomato flowers were collected in each sub-subplot on each of 13 dates in 2011 during the flowering period of the tomato crop, and the number of adult male and female F. occidentalis in each sample were determined (Figure 5). The effects of mulch, kaolin, and companion plant on each gender were analyzed using analysis of variance for a randomized complete block design for a split-split plot treatment arrangement for data across sample date using a mixed model (see Tyler-Julian et al.9 for a complete description of the analysis of variance and results). The main effects of mulch, kaolin, and companion plant were significant for the male western flower thrips (p < 0.01, 0.001, and 0.001, respectively), while the interactive effects of mulch X kaolin, mulch X companion plant, kaolin X companion plant, and mulch X kaolin X companion plant interactions were not significant (p > 0.05). These results showed that each of the main effects reduced the number of adult male F. occidentalis, and that the effects of each tactic were additive when combined with one another.
The main effect of mulch was significant for the female F. occidentalis (p < 0.01), while the main effects of kaolin and companion plants were not significant for the female F. occidentalis (p > 0.05). Therefore, the ultraviolet-reflective mulch reduced the female F. occidentalis in the tomato flowers, but kaolin and the companion plant did not. However, the mulch X kaolin interaction was significant (p < 0.05) showing that the combined effects of ultraviolet-reflective mulch and kaolin reduced the female F. occidentalis more than either tactic alone, while the kaolin applied to tomato on black mulch did not reduce female F. occidentalis numbers. The interactive effects of mulch X companion plant, kaolin X companion plant, and mulch X kaolin X companion plant interactions for female F. occidentalis were not significant (p > 0.05).
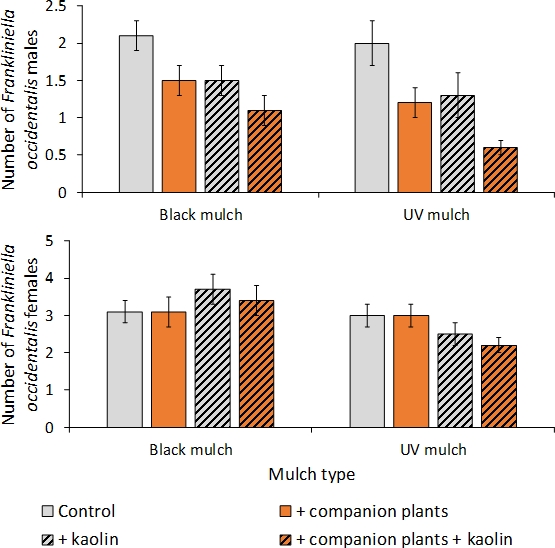
Figure 5: Example of analysis of data over sample date.
The mean number per 10 tomato flowers (SEM) of adult male and female F. occidentalis in mulch, kaolin, and companion plant treatments for sample data pooled across 13 dates in 2011 in a push-pull experiment conducted in Gadsden County, Florida. This figure has been modified from Tyler-Julian et al.9
The interaction of mulch X sample date was significant in the experiment in 2011 for male and female F. occidentalis adults (p < 0.01 and 0.001, respectively)9. This revealed that the ultraviolet-reflective mulch reduced flower thrips numbers on some, but not all, sample dates. Therefore, additional analyses were conducted to evaluate the effects of mulch on individual sample dates. The interaction showed that the ultraviolet-reflective mulch was effective in reducing flower thrips numbers early in the season, but there was no significance on individual sample dates during mid- or late-season (Figure 6).
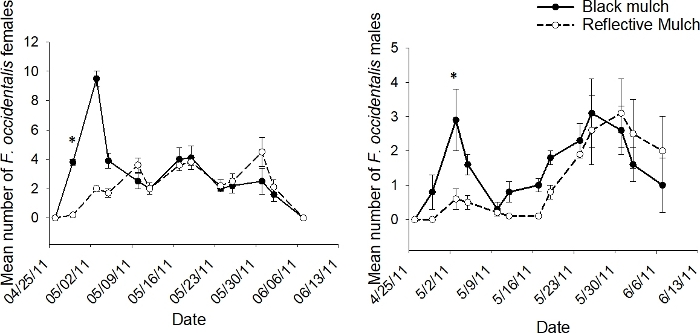
Figure 6: Example of population dynamics for whole plot treatment.
The mean number (+SEM) per 10 tomato flowers (n = 18 samples) of adult male and female F. occidentalis on each 2011 sample date in the whole plot treatment of black and ultraviolet-reflective mulch for data pooled across kaolin and companion plant treatments in the push-pull experiments conducted in Gadsden County, Florida (*indicates significance beyond 95% level of significance according to analysis of variance conducted for individual sample dates; d.f. = 1, 2). This figure has been modified from Tyler-Julian et al.9. Please click here to view a larger version of this figure.
The interaction of kaolin X sample date was not significant in 2011 for male or female F. occidentalis (p > 0.05)9. As previously shown above, the analyses of data pooled over sample date revealed that kaolin did not significantly affect female F. occidentalis numbers, while male F. occidentalis numbers were significantly reduced. The lack of a significant kaolin X sample date interaction in the analyses for data pooled over sample date suggested that the results for each gender were consistent across sample date (Figure 7).
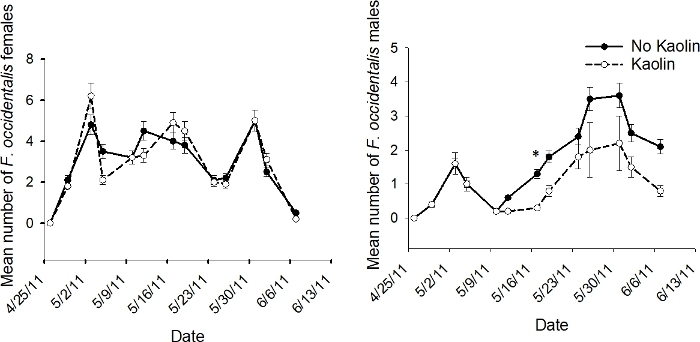
Figure 7: Example of population dynamics for subplot treatment.
The mean number (+SEM) per 10 tomato flowers (n = 12 samples) of adult male and female F. occidentalis on each 2011 sample date in the subplot treatment of kaolin and no kaolin for data pooled across companion plant treatments in the push-pull experiments conducted in Gadsden County, Florida (*indicates significance beyond 95% level of significance according to analysis of variance conducted for individual sample dates; d.f. = 1, 4). This figure has been modified from Tyler-Julian et al.9. Please click here to view a larger version of this figure.
The interaction of companion plant X sample date was significant in 2011 for male F. occidentalis (p < 0.05), but not for female F. occidentalis (p > 0.05)9. The analyses conducted to evaluate the effects of companion plant on individual sample dates revealed that companion plants reduced adult F. occidentalis numbers on late season sample dates, but never on early or mid-season sample dates (Figure 8).
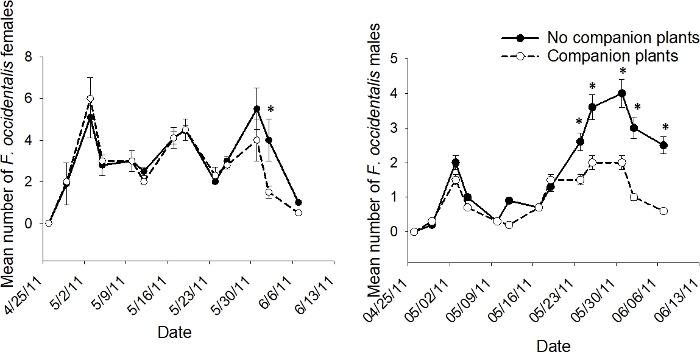
Figure 8: Example of population dynamics for sub-subplot treatment.
The mean number (+SEM) per 10 tomato flowers (n = 6 samples) of adult male and female F. occidentalis on each 2011 sample date in the sub-subplot treatment of companion plant and no companion plant in the push-pull experiments conducted in Gadsden County, Florida (*indicates significance beyond 95% level of significance according to analysis of variance conducted for individual sample dates; d.f. = 1, 8). This figure has been modified from Tyler-Julian et al.9. Please click here to view a larger version of this figure.
Data collected from the flowers of the companion plant in the study of Tyler-Julian et al.14 can be used to demonstrate the dynamic relationship between minute pirate bugs and its thrips prey in flowers (Figure 1B). As in the Tyler-Julian et al.9 study, the objectives were to determine the separate and combined effects of push factors (i.e., ultraviolet-reflective mulch and kaolin application) and a pull factor (i.e., the companion plant), on the population dynamics of Frankliniella species adult males and females in crop flowers. In the Tyler-Julian et al.14 study, the predominant flower thrips species was F. bispinosa in the companion plant H. annuus and in the pepper crop (>99% of the total thrips in the flowers). The thrips rapidly colonized the sunflowers and the pepper flowers, and their numbers were greatest soon after flowering began (Figure 9). Populations of thrips declined over time as the numbers of minute pirate bugs increased. The predator-prey ratio illustrated the ability of the predator to suppress the thrips populations with near extinction of the thrips populations occurring at ratios of >1 predator per 40 thrips.
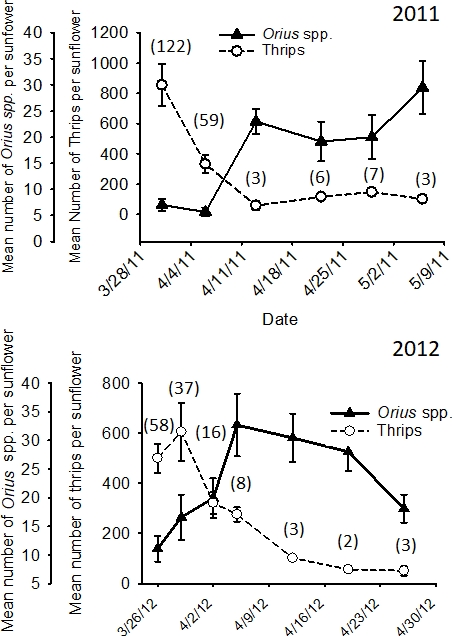
Figure 9: Example of evaluating the benefits of predation.
Mean number (+SEM) of total thrips (adults and larvae) and total Orius spp. (adults and nymphs) per Helianthus annuus flower head in experiments conducted in 2011 and 2012 in Palm Beach County, Florida (the number of total thrips prey per predator on each date shown in parenthesis). This figure has been adapted from data reported in Tyler-Julian et al.14 with permission from Oxford University Press.
