Generation and characterization of BIRFLU in vitro (Figure 1 and Figure 2)
A recombinant replication-competent IAV expressing two different reporter genes (BIRFLU) was constructed using state of the art molecular biology and plasmid-based reverse genetics techniques (Figure 1). Here, we chose to use Nluc due to several advantages over other luciferases, including its small size, ATP-independence, greater intensity, and optimized substrate48,60. Nluc was cloned into the HA segment of IAV PR8 followed by the porcine teschovirus (PTV) 2A cleavage site (2A) in front of the open reading frame (ORF) of HA (Figure 1). The ORF of HA included silent mutations to remove the original packing signals and avoid any possible recombination. The complete HA packaging signal was added in front of Nluc to allow proper incorporation of the modified HA segment into the virion and Nluc and HA expression from the same viral RNA segment (Figure 1). In addition, the fluorescent protein Venus was cloned into a modified IAV PR8 NS segment which encodes the two viral proteins NS1 and NEP from a single transcript32,36,41,54,57. To that end, Venus was fused to the C-terminal of NS1 and the entire NEP ORF was cloned downstream of the PTV 2A cleavage site that was placed between the NS1-Venus and NEP sequences (Figure 1). Ultimately, these two modified HA and NS viral plasmid constructs were used in combination with the rest of the IAV PR8 reverse genetics plasmids to generate BIRFLU (Figure 1). The in vitro and in vivo characterization of BIRFLU has been described previously55.
In Figure 2, we characterized in vitro BIRFLU by determining Venus, Nluc, and NP expression levels using fluorescence and indirect immunofluorescence approaches (Figure 2A,B). Confluent monolayers of MDCK cells were either mock-infected or infected (MOI 0.1) with WT or BIRFLU PR8 viruses and, at 18 h post-infection, Venus expression was directly evaluated using fluorescence microscopy (Figure 2A,B). Nluc (Figure 2A) and NP (Figure 2B) expression were visualized by indirect immunofluorescence using antibodies specific for each protein. As anticipated, Venus and Nluc expression were detected only in cells infected by BIRFLU and not in cells infected by WT PR8 virus. In addition, indirect immunofluorescence microscopy revealed NP expression in both WT and BIRFLU PR8-infected cells. No expression of Venus, Nluc or NP were detected, as expected, in mock-infected cells (Figure A,B).
To evaluate Nluc expression levels in vitro, MDCK cells were infected (MOI 0.001) with WT or BIRFLU PR8 viruses and Nluc activity in tissue culture supernatants was assessed at 24, 48, 72 and 96 h post-infection (Figure 2C). Only Nluc activity was detected in tissue culture supernatants of MDCK cells infected with BIRFLU (Figure 2C). Nluc activity in the tissue culture supernatants was detected as early as 24 h post-infection with higher expression levels at 96 h post-infection, most probably because the cytopathic effect (CPE) induced during viral infection release the Nluc protein retained into the cell. To evaluate the fitness of BIRFLU in cultured cells, growth kinetics of WT and BIRFLU PR8 viruses were also evaluated (Figure 2D) and the presence of infectious virus in tissue culture supernatants was determined by immune-focus assay (Figure 2D). Notably, BIRFLU replication kinetics were comparable to those of WT PR8 virus, although BIRFLU replication was slightly delayed and did not reach same viral titers as WT PR8. However, BIRFLU was able to reach titers of 5 x 107 PFU/mL (Figure 2D), indicating that expression of two reporter genes in the viral genome does not significantly interfere with BIRFLU replication in MDCK cells.
Tracking BIRFLU infection in mice (Figure 3 and Figure 4)
Figure 3 is a schematic flow chart for the assessment of BIRFLU dynamics in a mouse model of IAV infection. Five-to-seven-week-old female BALB/C mice were either mock-infected with 1x PBS or infected with 1 x 106 PFU of BIRFLU intranasally. At 3 days post-infection, mice were anesthetized with isoflurane and then injected with Nluc substrate retro-orbitally. All mice were immediately placed in the IVIS instrument and Nluc signal was assessed in vivo using the IVIS. Next, mice were euthanized and lungs were harvested. Excised lungs were then analyzed ex vivo using the in vivo imager to determine fluorescence intensity via Venus expression. Lastly, mice lungs were homogenized, and viral titers and stability were determined by plaque assay. Plaques were assessed by the direct fluorescence of Venus, by immunostaining using antibodies specific for Nluc and by crystal violet staining.
Previously described replication-competent reporter-expressing IAVs express a single reporter gene, most frequently either a fluorescent or a bioluminescent protein, as surrogate for viral infection and replication. However, BIRFLU, is able to express both types of reporter genes upon viral infection. To assess the correlation between bioluminescence (in vivo imaging) and fluorescence (ex vivo imaging) after BIRFLU infection, five-to-seven-week-old female BALB/c mice were mock-infected with 1x PBS or inoculated with BIRFLU (106 PFU) intranasally. Nluc activity (Figure 4A) was evaluated by administration of Nluc substrate injected retro-orbitally at 3 days post-infection using an in vivo imaging instrument. We chose to evaluate bioluminescence at day 3 because previous studies indicated that IAV replication, including PR8, peaks between days 2 and 4 post-infection24,54. Bioluminescence was monitored (Figure 4A, top) and used to calculate the average total flux (Flux (log10 p/s) (Figure 4A, bottom). As predicted, mice inoculated with BIRFLU displayed high bioluminescence activity but no signal was detected in mock-infected mice. Thereafter, the lungs of infected mice were harvested and Venus expression was assessed using ex vivo imaging (Figure 4B, top). Moreover, the fluorescence average radiant efficiency was calculated (Figure 4B, bottom). The excised mice lungs were also homogenized to determine the viral titers and the genetic stability of BIRFLU in vivo (Figure 4C,D). Genetic stability of BIRFLU was analyzed through plaque assay using the viruses isolated from mice lungs and fluorescent microscopy (Venus, top), immunostaining (Nluc, middle) and crystal violet staining (bottom). BIRFLU recovered from mice lungs were able to form plaques and stably expresses both reporter genes (Figure 4C). Notably, we observed a good correlation between bioluminescence and fluorescence signals with viral replication.
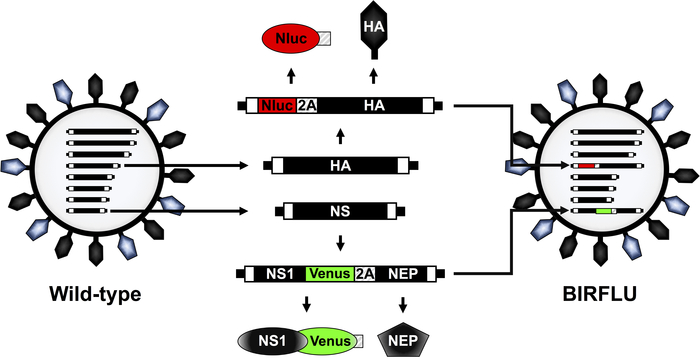
Figure 1: Schematic representation of IAV PR8 WT and BIRFLU virion structure and genome segments. IAV are surrounded by a lipid bilayer containing the two major viral glycoproteins hemagglutinin (HA; black) and neuraminidase (NA; blue). IAV contain eight single-stranded, negative-sense, RNA segments (PB2, PB1, PA, HA, NP, NA, M, and NS). Each viral segment contains non-coding regions (NCR) at the 3’ and 5’ ends (black boxes). Also, at the 3’ and 5’ end of the viral (v)RNAs are the packaging signals, responsible for the efficient encapsidation of vRNAs into nascent virions (white boxes). IAV PR8 HA and NS viral segments and products are indicated in black. Sequences of Nluc, Venus, and PTV 2A are indicated in red, green and striped boxes, respectively. The schematic representation of the modified HA and NS segments expressing Nluc and Venus, respectively, in BIRFLU are also indicated. This figure has been adapted from Nogales et al.55. Please click here to view a larger version of this figure.
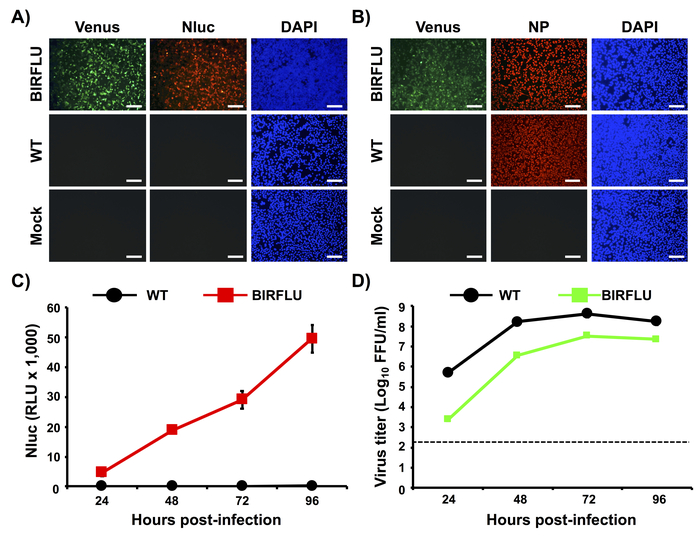
Figure 2: In vitro characterization of BIRFLU. (A, B) Analysis of protein expression by direct fluorescence and immunofluorescence. MDCK cells were mock-infected or infected (MOI 0.1) with PR8 WT or BIRFLU viruses. Infected cells were fixed at 18 h post-infection to directly visualize Venus expression by direct fluorescent microscopy and to visualize Nluc (A) and viral NP (B) expression using specific antibodies and indirect immunofluorescence. Nuclei were stained with DAPI. Representative images (20x magnification) are shown. Scale bars = 100 μm. (C, D) Growth kinetics of PR8 WT and BIRFLU. Nluc activity (C) and viral titers (D) in tissue culture supernatants from MDCK cells infected (MOI 0.001) with WT and BIRFLU PR8 viruses were assessed at the indicated times post-infection. Data represent the means ± SD of triplicates. Viral titers were determined by immune-focus assay (FFU/mL). Dotted line denotes the limit of detection (200 FFU/ml). This figure has been adapted from Nogales et al.55. Please click here to view a larger version of this figure.
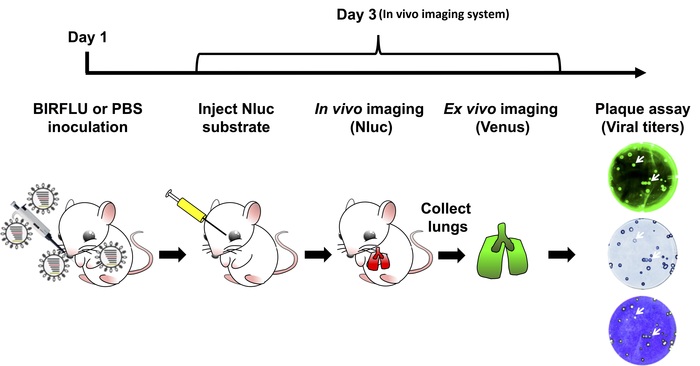
Figure 3: Schematic representation for the study of BIRFLU in mice. Expression of Nluc and Venus reporter genes was evaluated in mice infected with 1 x 106 PFU of BIRFLU using in vivo or ex vivo imaging. Briefly, on day 1, 5 to 7 week-old female BALB/c mice were mock-infected (1x PBS) or inoculated with 1 x 106 PFU of BIRFLU intranasally. At day 3 post-infection, mice were mildly anesthetized using isoflurane and Nluc substrate was injected retro-orbitally. Nluc signal was directly assessed using in vivo imaging. Immediately after imaging, mice were euthanized and expression of Venus in whole excised lungs was analyzed using ex vivo imaging. Recovered mice lungs were homogenized to evaluate viral replication and stability by plaque assay. Arrows indicate correlation between fluorescence (Venus), immunostaining (Nluc) and crystal violet staining. Please click here to view a larger version of this figure.
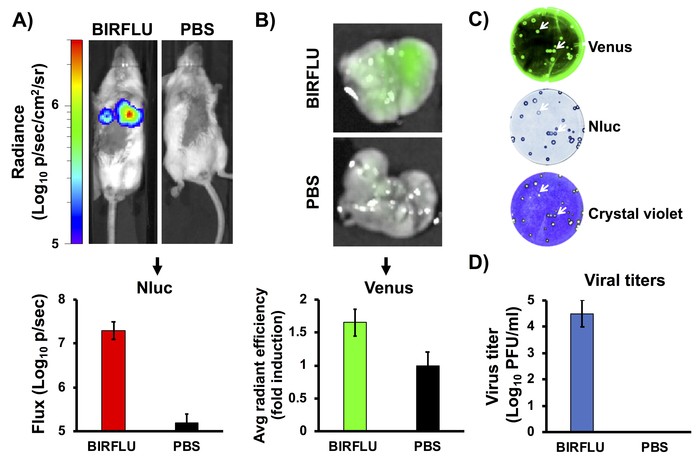
Figure 4: In vivo bioluminescence and fluorescence expression. Female five-to-seven-week-old BALB/c mice were mock-infected (1x PBS) or inoculated with 1 x 106 PFU of BIRFLU intranasally. At day 3 post-infection, Nluc activity (A) in the whole mouse was determined. Representative images of a single mouse showing radiance scale (p/sec/cm2/sr). Bioluminescence radiance values were quantitated and the average total flux is shown (Flux (Log10 p/s). After Nluc imaging, lungs were harvested for ex vivo imaging (B). Representative pictures from whole lungs are shown. To quantify Venus expression, mean values of regions of interest (ROIs) were normalized to lung auto-fluorescence from mock-infected mice and fold changes were calculated. To analyze the genetic stability of BIRFLU in vivo, viruses recovered from mice lungs were analyzed by plaque assay using fluorescent microscopy (Venus, top), immunostaining (Nluc, middle) and crystal violet staining (bottom) (C). Representative images from one mouse are shown. To evaluate virus replication, whole lungs were homogenized after imaging and used to infect MDCK cells and determine viral titers by plaque assay (PFU/mL) (D). Arrows indicate correlation between fluorescence (Venus), immunostaining (Nluc), and crystal violet staining. Bars represent the mean ± SD of lung virus titers. This figure has been adapted from Logales et al.55. Please click here to view a larger version of this figure.
| Tissue culture media and solutions | Composition | Storage | Use |
| Tissue culture media: Dulbecco’s modified Eagle’s medium (DMEM), 10 % Fetal Bovine Serum (FBS), 1% Penicillin-Streptomycin-L-glutamine (PSG) (DMEM 10 % FBS 1% PSG). | 445 ml DMEM, 50 mL of FBS and 5 mL of 100x PSG. | 4 °C | Maintenance of MDCK cells |
| Post-infection media: DMEM 0.3% Bovine Albumin (BA), 1% PSG (DMEM 0.3 % BA 1% PSG). | 491 ml DMEM, 4.2 ml of 35 % BA and 5 ml of 100x PSG | 4 °C | Maintenance of MDCK cells after viral infection |
| 10x Phosphate buffered saline (PBS) | 80 g of NaCl, 2 g of KCl, 11.5 g of Na2HPO4.7H2O, 2 g of KH2PO4. Add ddH2O up to 1 L. Adjust pH to 7.3 | Room temperature | To prepare 1x PBS |
| 1x PBS | Dilute 10x PBS with ddH2O | Room temperature | Wash cells |
| Infection media: 1x PBS, 0.3% BA, 1% Penicillin-Streptomycin (PS) (PBS/BA/PS). | 487 mL 1x PBS sterile, 4.2 mL of 35% BA and 5 ml of 100x 1% PS(100 U/mL) | 4 °C | Viral infections |
| Fixation/permeabilization solution: 4% formaldehyde, 0.5% triton X-100 diluted in 1x PBS. | 400 mL neutral buffered formalin 10%, 5 ml of Triton X-100 and 595 mL of 1x PBS | Room temperature | Fix and permeabilization of MDCK cells. |
| Blocking solution: 2.5% Bovine Serum Albumin (BSA) in 1x PBS. | 2.5 g of BSA in 97.5 mL of 1x PBS | 4 °C | Blocking solution for immunofluorescence and plaque assays. |
| Antibody dilution solution (1% BSA in 1x PBS) | 1 g of BSA in 99 mL of 1x PBS | 4 °C | Dilution of primary and secondary antibodies. |
| 0.1% crystal violet solution | 1 g of crystal violet in 400 mL of methanol. Add 600 ml of ddH2O | Room temperature | Staining of MDCK cells in plaque assays. |
| Tosylsulfonyl phenylalanyl chloromethyl ketone (TPCK)-treated trypsin | Prepare a 1,000x stock solution at 1 mg/mL in ddH2O | -20 °C | For viral infections. |
Table 1: Tissue culture media and solutions.
