All experimental protocols described were approved by the University of Sydney Animal Ethics Committee (approved protocol number: 2018/1308).
1. Animals
NOTE: Mice were obtained from the Australian Rodent Centre (ARC; Perth, Australia) and held at the Medical Foundation Building Animal Facility at the University of Sydney.
- Maintain the mice on a normal 12 h light/dark cycle with environmental enrichment.
- Use male and female C57BL/6 mice (3–5 weeks old) for all experiments.
2. Preparation of Solutions
- Prepare 1 L of artificial cerebrospinal fluid (ACSF) composed of 29 mM NaHCO3, 11 mM glucose, 120 mM NaCl, 3.3 mM KCl, 1.4 mM NaH2PO4, 2.2 mM MgCl2, 2.77 mM CaCl2.
- Prepare 200 mL of sucrose-ACSF (sACSF) containing 29 mM NaHCO3, 11 mM glucose, 241.5 mM sucrose, 3.3 mM KCl, 1.4 mM NaH2PO4, 2.2 mM MgCl2, 2.77 mM CaCl2. Prior to the inclusion of CaCl2 to the ACSF and sACSF, gas the solutions with carbogen (95 % O2 and 5 % CO2) to establish a pH of 7.4 and avoid calcium precipitation (cloudiness).
- Prepare K+-based intracellular solution composed of 70 mM potassium gluconate, 70 mM KCl, 2 mM NaCl, 10 mM HEPES, 4 mM EGTA, 4 mM Mg2-ATP, 0.3 mM Na3-GTP; with a final pH of 7.3 (adjusted using KOH).
NOTE: It is recommended to filter intracellular solutions with 0.22 µm filters and store 0.5 mL aliquots of the solution at -20 °C.
3. Preparation of the Brainstem
- Prior to brainstem extraction, equilibrate the sACSF with carbogen and cool at -80 °C for 25 min so that an ice slurry is formed.
- Anaesthetize the mouse with isoflurane (3–5 %) saturated in oxygen (3 mL/min). Once the hind paw reflexes are absent, decapitate the mouse with sharp stainless-steel scissors.
- Expose the skull by making a sagittal incision in the skin using a razor blade (#22 rounded).
- Using the pointed end of a pair of standard pattern scissors make a small incision at the lambda and cut along the longitudinal fissure.
- Carefully reflect away the paired parietal bones and the occipital bones using a pair of shallow-bend Pearson rongeurs.
NOTE: During this whole procedure the brain is continuously bathed in situ using the previously prepared ice-cold sACSF slurry. - Isolate the brainstem from the forebrain and its bony encasing using a razor blade (#11 straight) to cut down the parieto-occipital sulcus and at the caudal medulla.
- Mount the isolated brainstem ventral end down on a previously cut trapezoidal polystyrene block. Remove excess fluid around the dissected tissue with a wick of tissue paper to ensure good tissue adhesion to the cutting stage.
NOTE: The polystyrene block is cut in a trapezoidal shape, to ensure the rostral end of the midbrain fits and tapers into the spinal cord. - Use cyanoacrylate glue to fix the polystyrene block with the attached brainstem rostral end down to the cutting stage.
- Using an advance speed of 0.16 mm/s and vibration amplitude of 3.00 mm, prepare 200 µm transverse slices of the MVN.
NOTE: Location of the MVN is determined using the Paxinos and Franklin mouse brain atlas (Figures 79–89)20. The MVN (listed as MVe in atlas) lies immediately ventrolateral to the 4th ventricle and is largest right before the attachment of the cerebellum (between the inferior colliculi and the obex). - Use a plastic-trimmed pipette to transfer slices onto a filter paper disc sitting in carbogenated ACSF at 25 °C for at least 30 min prior to recording.
4. Instruments
- Use a standard electrophysiological setup to perform whole-cell patch clamp techniques21.
- Prepare micropipettes using a two-step protocol (heat step 1: 70; heat step 2: 45) on a micropipette puller (see the Table of Materials). Micropipettes should have a final resistance ranging 3–5 MΩ with internal solution when placed in the bath.
NOTE: Settings used may vary depending on the temperature within room and can change quite frequently.
5. Whole-cell Patch Clamp Electrophysiology
- To obtain whole-cell patch clamp recordings from individual neurons in the MVN, a K+-based internal solution is used within the recording pipette.
- Transfer a single tissue slice from the incubation chamber to the recording chamber and secure the slice using a nylon thread on a U-shaped weight. Continuously perfuse the recording chamber with carbogenated-ACSF at 25 °C at a flow rate of 3 mL/min.
- After filling a micropipette with internal solution, locate the MVN using a low power (10x) objective lens. Using a high-power (40x) objective, individual neurons within the MVN can be located.
NOTE: Cell quality is essential in ensuring quality recordings and durability of the cell when attempting to achieve the whole-cell configuration. A good cell will demonstrate spherical shape, a reflective cell membrane and an invisible nucleus. A bad cell will have a large visible nucleus (egg-like) and a swollen/shrunken appearance. - Before breaching the tissue with the pipette, apply a small amount of positive pressure to push debris away from the pipette tip.
- Move the pipette using the micromanipulator towards the chosen neuron and a small dimple should form on the neuronal membrane. Release positive pressure and apply a small amount of negative pressure.
- Once a 1 GΩ seal is achieved, apply gentle short and sharp positive pressure to the pipette holder through the suction port to rupture the membrane and create a whole-cell configuration.
- Make whole-cell current clamp recordings using standard techniques21,22.
6. Applying Sinusoidal and Stochastic Noise to Individual Medial Vestibular Nucleus Neurons
- Apply the stochastic and sinusoidal noise at a range of amplitudes from 3 to 24 pA to determine neuronal threshold and firing rate.
- Determine the sensory threshold by grouping lower and higher stimulus intensities and perform an ANOVA to observe any differences (as shown in Supplementary Figure 1).
- Calculate the average firing rate over the 10 s period where the depolarizing current step was/will be injected for each individual current level (i.e., 7 total episodes; Figure 1).
- Use the average firing rate values to generate a firing rate versus current plot and perform a linear regression analysis to determine the gradient of the line of best fit. The gradient of the line of best fit is indicative of the neuronal gain22.
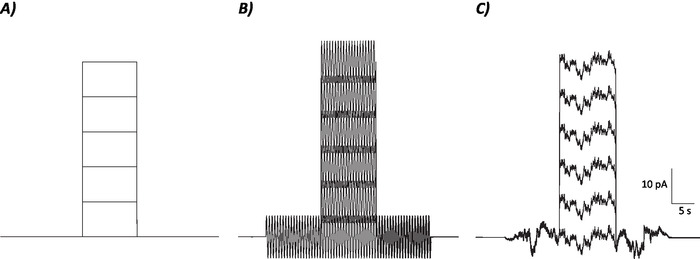
Figure 1: Diagrammatic profiles of control, sinusoidal and stochastic noise protocols. (A) Control (no noise) protocols applied to MVN neurons. (B) Sinusoidal noise protocol with a frequency of 2 Hz. (C) Stochastic noise protocols where majority of the power spectrum is ≤2 Hz. Each protocol presented here has an amplitude of ±6 pA with a 10 s depolarizing current increasing by 10 pA up to 50 pA. The true stimulus does not have a depolarizing current step and is therefore the first episode of these protocols to determine neuronal gain changes. Please click here to view a larger version of this figure.
Initial recordings can provide information about the effects that sinusoidal and stochastic noise have on basal firing rates of individual MVN neurons and how the stimuli effect the gain of neurons. Figure 2 shows that neither sinusoidal nor stochastic noise change basal firing rates of MVN neurons when compared to control (no noise) recordings. This information is crucial for determining the threshold of the individual neurons. During the application of galvanic vestibular stimulation to humans, a sensory thresholding task is performed to ensure that the stimulus is subthreshold13. The subthreshold stimulus is an important component of the stochastic resonance (SR) phenomenon7,8. In vitro, this thresholding task needs to be performed differently and the activity or basal firing rate of neurons has been chosen for this. This ensures that the stimuli are as close to subthreshold as possible and therefore comparable to human studies. Figure 2B highlights that the selected noise level (6 pA) is subthreshold, as it can be observed that average firing rate begins to increase from 12 pA (experimental threshold). This threshold was determined objectively by grouping stimulus levels above (18 and 24 pA) and below (3 and 6 pA) the 12 pA threshold and is shown in Supplementary Figure 1.
Next, neuronal gain was evaluated by subjecting neurons to a suite of depolarizing current steps (0-50 pA, increasing by 10 pA) with and without (control) noise (Figure 1). These results are crucial to determining the effect that stochastic noise may have on neurons in the central vestibular system and thus, potentially how GVS is eliciting its effects on human balance. Figure 3 shows that sinusoidal (Figure 3B) and stochastic (Figure 3A) noise applied at subthreshold amplitudes of 6 pA can alter the gain of MVN neurons. These results were assessed by measuring the firing rate during each 10 s current step and performing a linear regression analysis to calculate the gain (gradient) from the line of best fit.
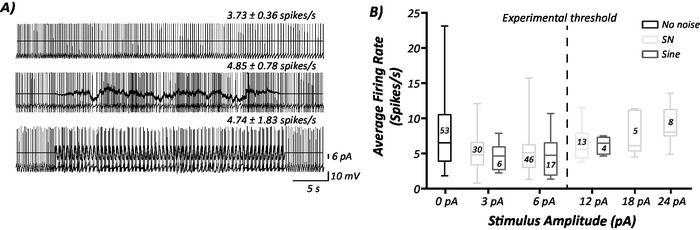
Figure 2: The effect of sinusoidal and stochastic noise on MVN neuronal firing rate. (A) Stochastic (SN; middle trace) and sinusoidal noise (bottom trace) at a 6 pA amplitude show no significant effect on basal firing rate of an individual MVN neuron in comparison to control (no noise; top trace). (B) Firing rate of MVN neurons in response to control (n = 53), stochastic and sinusoidal noise protocols (with no current steps) of amplitudes 3 (SN, n = 30; sine, n = 6), 6 (SN, n = 46; sine, n = 17), 12 (SN, n = 13; sine, n = 4), 18 (SN, n = 5; sine, n = 0) and 24 (SN, n = 8; sine, n = 0) pA. Lines/whiskers indicate the maximum and minimum values, the box indicates the 25th-75th percentiles and the line within the box indicates the mean firing rate (spikes/s). The dashed line indicates experimental threshold, as chosen by pooling the mean firing rates within 3 and 6 pA (below 12 pA) and 18 and 24 pA (above 12 pA) shown in Supplementary Figure 1. Please click here to view a larger version of this figure.
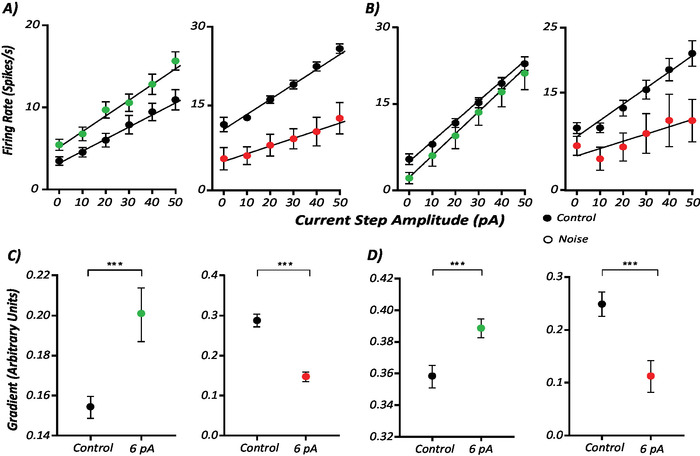
Figure 3: Sinusoidal and stochastic noise alter MVN neuronal gain. (A) MVN neuronal firing rate at each depolarizing current step and the corresponding gain calculation in response to stochastic noise. (B) The data presented were generated the same way as in Figure 3A but during the application of sinusoidal noise. (C,D) Graphs represent the gains calculated from the lines of best fit of A and B. Error bars indicate S.D. Statistical significance was determined by linear regression analysis comparing the gradients of the lines of best fit between control and experimental condition. **p < 0.02; ***p < 0.01. Please click here to view a larger version of this figure.
Supplementary Figure 1: Objective determination of 12 pA threshold. Firing rates for less than 12 pA (3 and 6 pA) and more than 12 pA (18 and 24 pA) were pooled and averaged. These averages were then analyzed using an ANOVA and statistical significance between sham and >12 pA and between <12 pA and >12 pA. *p < 0.05. Please click here to download this file.
