Parameters of action potentials and several membrane properties can be calculated on the basis of intracellular recordings when stable conditions of cell penetration are ensured. Figure 1A presents a typical orthodromic action potential evoked by intracellular stimulation, which meets all criteria for data inclusion (the resting membrane potential of at least -50 mV, and spike amplitude higher than 50 mV, with a positive overshoot). Action potential parameters, such as the spike amplitude, the afterhyperpolarization amplitude or the afterhyperpolarization half-decay time (AHP-HDT) can be measured. A value of the latter parameter in rat motoneurons serves as a reliable criterion for distinguishing between fast and slow motoneurons (AHP-HDT > 20 ms for slow, while AHP-HDT <20 ms for fast motoneurons)17. Figure 1B shows a cell response to a 100 ms hyperpolarizing current pulse of 1nA, from which both peak and plateau input resistance (IR) of a motoneuron can be determined from the voltage deflection. Figure 1C shows an expanded voltage trace of a rheobasic spike with a clearly marked voltage threshold of the spike, indicating the level of membrane depolarization at which voltage-gated sodium channels are activated to initiate the action potential. All these recordings can be repeated several times during and after tsDCS application, which allows us compare respective parameters as long as the resting membrane potential is stable and other criteria of stimulation and recording protocol are fulfilled.
Several studies have indirectly shown that tsDCS alters motoneuron excitability and firing pattern9,18. Figure 2 shows examples of intracellular voltage traces from two motoneurons stimulated intracellularly with 500 ms square pulses of depolarizing current before, during and after tsDCS application. Under stable conditions, recordings repeated several minutes one after another can be performed, and motoneuron firing patterns can be reliably compared. Anodal (+) tsDCS was found to act towards increased motoneuron excitability and higher frequencies of rhythmic firing (Figure 2A), while cathodal (-) tsDCS acted towards firing inhibition (Figure 2B). Moreover, the effects of both types of tsDCS outlasted the period of polarization. It is also worth noting that the observed changes in excitability and firing pattern are not merely a result of cell membrane depolarization or hyperpolarization by anodal or cathodal tsDCS, respectively, but display profound alterations not related to the change of a membrane potential, as they persisted despite the fact that this parameter returned to a baseline after the end of polarization.
Finally, it has to be stressed that any deviations from the presented protocol will likely result in a failed experiment, due to deterioration of preparation and/or a profound decline of data reliability. Figure 3 shows examples of recordings when data inclusion criteria were compromised either due to imperfect cell penetration (Figure 3A), neglection to compensate microelectrode resistance and capacitance (Figure 3B) or a spinal cord instability (Figure 3C). It is important that researchers identify such non-optimal recordings, and implement proper corrective actions or disregard such results from the data set.
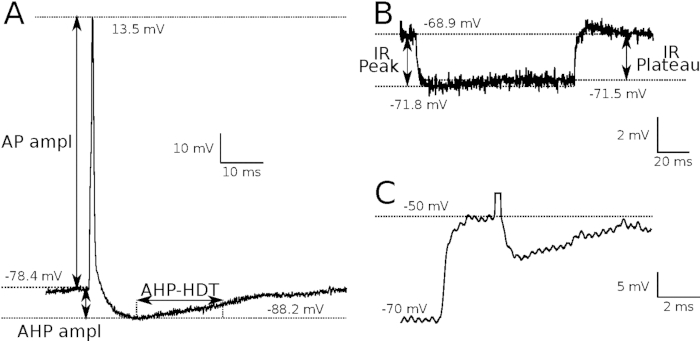
Figure 1: Parameters of action potentials and membrane properties.
(A) An orthodromic action potential elicited by intracellular stimulation, with indicated basic parameters which can be calculated from this record. AP ampl = action potential amplitude; AHP ampl = afterhyperpolarization amplitude; AHP-HDT = afterhyperpolarization half-decay time. (B) The voltage trace of a membrane response to a short (100 ms) depolarizing current pulse of 1nA intensity, which enables us to calculate input resistance (IR). Notice the peak of a potential deflection (IR Peak) followed by a small decrease and the following plateau phase of the membrane potential (IR plateau). (C) The expanded voltage trace of a rheobasic spike with a dotted horizontal line indicating the spike voltage threshold. Please click here to view a larger version of this figure.
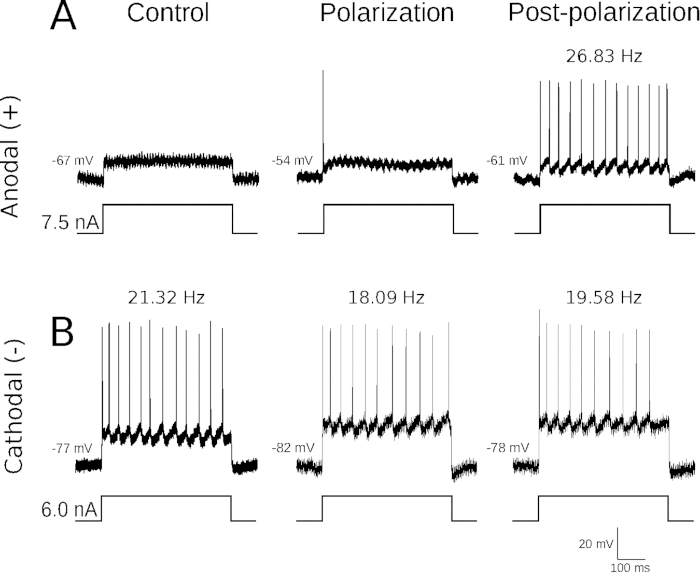
Figure 2: Effects of polarization on motoneuron firing.
(A) Intracellular records from one motoneuron stimulated intracellularly with 7.5 nA for 500 ms, made before (left), during anodal tsDCS (0.1 mA, middle), and 10 min after the end of polarization (right). Note the gradual increase in the motoneuron excitability at the same stimulus intensity. (B) Intracellular records from another motoneuron stimulated intracellularly with 6 nA for 500 ms, made before (left), during cathodal tsDCS (0.1 mA, middle), and 10 min after the end of polarization (right). Note a gradual inhibition of motoneuron firing frequency at the same stimulus intensity. Below recordings, traces of intracellular stimulation current are provided. The calibration bars in the bottom right apply to all presented intracellular recordings. The values of the resting membrane potential are provided to the left of each recording. Frequencies of steady-state firing, calculated from the means of the final three interspike intervals, are given above records. Please click here to view a larger version of this figure.
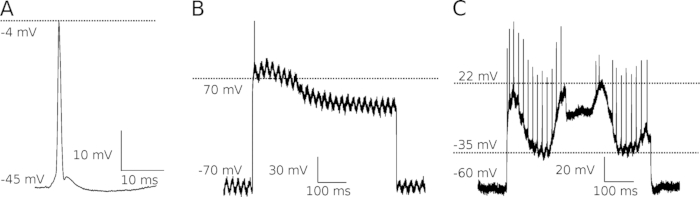
Figure 3: Examples of suboptimal records as a result of deviations from the experimental protocol.
(A) The antidromic spike recorded from a motoneuron inadequately penetrated. The resting membrane potential is insufficient (-45 mV), and despite an appropriate shape of the spike with all consecutive phases of depolarization, repolarization, and hyperpolarization, its amplitude is too low (41 mV) and without an overshoot. (B) A rheobasic spike generated at an unrealistic voltage threshold (membrane depolarized to +68 mV). This kind of error is usually due to a blocked microelectrode, with uncompensated resistance and capacitance. One can also see that this record is strongly contaminated by 50 Hz electrical noise. (C) A motoneuron rhythmic firing in response to 500 ms depolarizing current, with large fluctuations of a membrane potential, predominantly caused by unstable microelectrode penetration, possibly due to excessive respiratory movements. For all the presented cases the calculated membrane or firing properties would be unreliable. Please click here to view a larger version of this figure.
