A representative result of this sorting protocol is shown in Figure 3. The total sorting time of two testes (one mouse) is usually around 3 hours, which is dependent on the concentration of cell suspension and the sorting speed. After sorting, the purity of spermatocytes is confirmed by immunostaining of SYCP3 and γH2AX (Figure 3A). The representative purity of sorted L/Z, P, D spermatocyte fractions are around 80.4%, 90.6%, and 87.6%, respectively (Figure 3C). We have determined the substages based on the criteria we published previously19. Briefly, in the leptotene and zygotene stage, synapsis between homologous chromosomes is incomplete, which is indicated by thin threads of SYCP3 staining. Broad γH2AX domains are observed throughout the nuclear chromatin due to programmed DNA double-strand breaks. In the pachytene stage, homologous chromosomes have completely synapsed, and γH2AX specifically accumulates on the sex chromosomes. In the diplotene stage, homologous chromosomes progressively undergo desynapsis. The purity of RS is confirmed by nucleus staining with DCV (Figure 3B). RS can be precisely judged with DNA staining: a unique DAPI-intense chromocenter surrounded by euchromatin; or combine with specific markers, such as Sp56 that is expressed within the developing acrosomal granule of spermatid and histone variant H1T that is highly expressed in nucleus after the mid-pachytene stage (Figure 3B).
RS purity is around 90.1% after sorting (Figure 3C). The sample size of purity analysis is over 1,000 cells for each experiment; the purity of L/P and D populations is averaged from 6 independent experiments; the purity of P and RS populations is averaged from 3 independent experiments. The viability of these isolated cells is usually over 95% (Figure S1). The total yield of each fraction from a single adult mouse is estimated and listed in Fig 3C, which provides sufficient cells for various downstream analyses. Recently, we have used this protocol to isolate wild-type pachytene spermatocytes for ChIP-seq analysis20,21.
| Reagent | Ingredient | Stock concentration | Volume |
| (HBSS base) | |||
| Dissociation Buffer | DMEM | – | 2 ml |
| (DMEM base) | FBS | – | 40 μl |
| Hyaluronidase | 100 mg/ml | 30 μl | |
| DNase I | 10 mg/ml | 50 μl | |
| Collagenase Type I | 100 mg/ml | 40 μl | |
| Recombinant Collagenase | 14000 unit/ml | 100 μl | |
| FACS buffer | PBS | – | 980 ml |
| (PBS base) | FBS | – | 20 ml |
Table 1: Reagent Recipe. The dissociation buffer must be prepared right before use. Prewarm DMEM before starting dissection. The enzyme stocks can be prepared any time before the experiment and stored at -20 °C. FACS buffer needs to be vacuum-filtered and stored at 4 °C; prewarm to room temperature before use.
Figure 1: Workflow of murine spermatogenic cells isolation on DCV-based sorting. This image illustrates the general procedure, from tissue dissociation to FACS sorting, to harvesting of isolated spermatogenic cells within one day. Please click here to view a larger version of this figure.
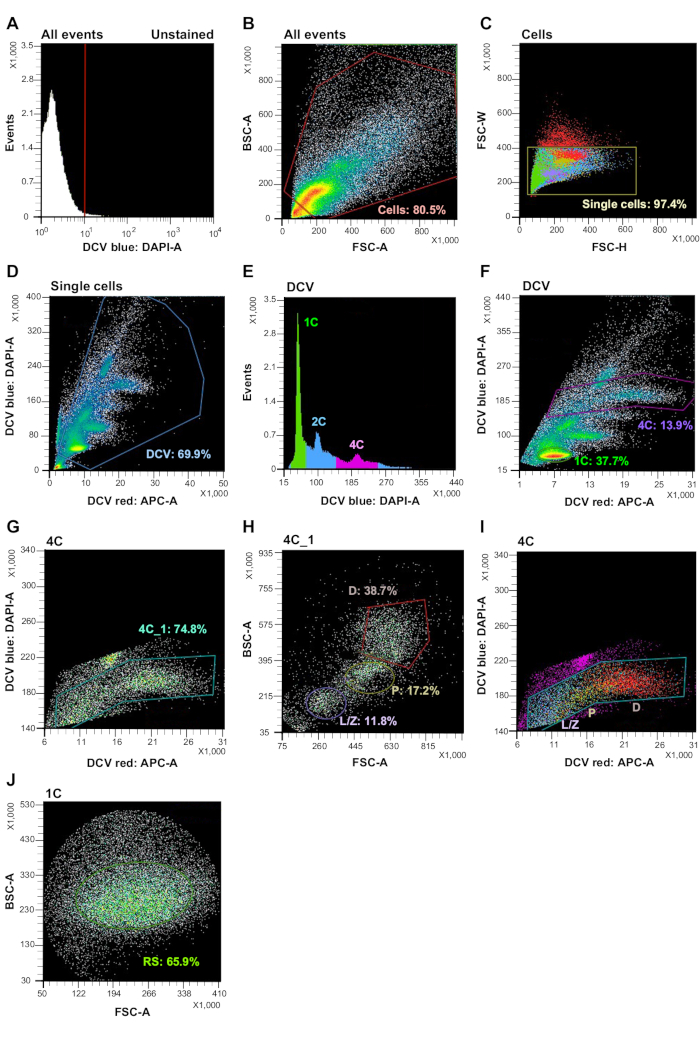
Figure 2: FACS analysis of adult murine testicular cells based on DCV fluorescence and light scattering. (A) Acquired unstained cells in the first decade of a DCV-blue histogram plot (left side of the red bar). (B)(C) Debris and non-single cell excluded by light scattering. (D) Unstained cell and side population exclusion based on DCV fluorescence. (E) DNA content determination based on DCV-blue fluorescence. Left peak (green) and right peak (pink) correspond to 1C and 4C populations. (F) Gating on 1C and 4C testicular populations based on DCV-blue/DCV-red fluorescence. (G) Precise gating on 4C testicular populations. (H) Back-gating of Gate 4C from the DCV plot on an FSC/BSC plot. Based on regions of minimal overlap on the FSC/BSC plot, the L/Z, P, and D gates are created to enrich their respective spermatocyte populations. (I) Color dot plot showing the L/Z, P, and D populations are in continuous order within Gate 4C. (J) Back-gating of Gate 1C from the DCV plot on an FSC/BSC plot. RS gate was created to enrich round spermatid population with uniform size, resulting in greater purity of populations during sorting. Please click here to view a larger version of this figure.
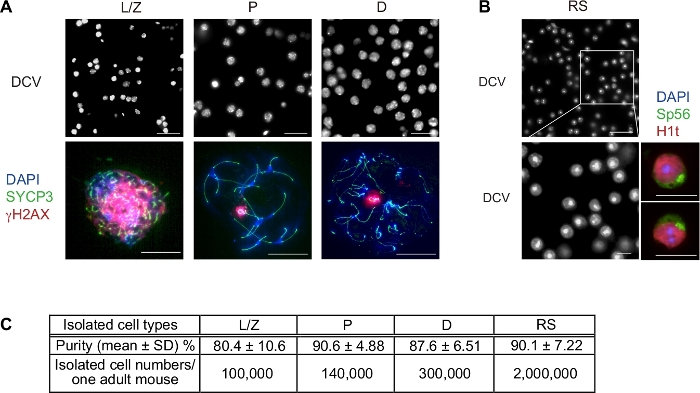
Figure 3: Representative result images and statistics of spermatogenic cells obtained from sorting. (A) Immunofluorescence characterization of sorted spermatocytes. Upper panel: DCV staining showing nucleus pattern of the live spermatocytes right after sorting; L/Z (leptotene/zygotene), P (pachytene), and D (diplotene). Lower panel: Confirmation of meiotic substages for each population by immunostaining for SYCP3 (green) and γH2AX (red). (B) Representative DCV image showing nucleus pattern of RS. Scale bars: 50 μm (upper panels), and 10 μm (lower magnified panels). Right Panels: Immunofluorescence confirmation of round spermatids stained with Sp56 and H1T. (C) The purity of L/Z, P, and D were confirmed by immunostaining, sample size was over 1,000 cells for each independent experiment, in total 6 independent experiments. The RS purity was confirmed by nucleus staining with a total of 3 independent experiments. The total cell number of the testicular cell suspension from one 8-week-old WT B6 mouse was around 100 million cells before sorting. Please click here to view a larger version of this figure.
Figure S1: The viability of isolated pachytene spermatocytes. A representative image shows the cell viability of isolated pachytene spermatocytes (Red: PI; Blue: DCV). PI could not be combined with DCV during sorting. However, under microscope, the DCV-red signal was quite low; therefore, PI-positive dead cells were easily distinguished from other live cells. The viability is usually over 95%. Scale bars: 10 μm. Please click here to download this figure.
Figure S2: Incomplete dissociation or debris disturbs gating. The A population (red circle) contains debris and polymer of spermatids (indicated by arrow). The bigger A population will cross with B population (yellow circle) and eventually contaminate the 4C population. Scale bars: 200 μm. Please click here to download this figure.
