The following protocols describe analysis performed on human sputum. Human sample handling was approved by the ethics committee of the University of Heidelberg and written informed consent was obtained from all patients or their parents/legal guardians (S-370/2011) and healthy controls (S-046/2009).
NOTE: The following protocols describe the sample preparation and the quantification of neutrophil serine proteases (NSPs) activity. The experimental procedures presented herein focus on human sputum and neutrophil elastase14,20,21 (NE) or cathepsin G15 (CG) activity measurement. However, slight adaptations in the sample preparation protocol render the analysis of blood-derived cells and tumor homogenates possible. In addition, matrix metalloproteinase 12 and cathepsin S activities can be investigated similarly by means of dedicated FRET probes22,23,24,25,26.
1. Sample preparation: cell isolation and supernatant separation
NOTE: If possible, the treatment of sputum should be carried out within 120 min after expectoration and the sputum should be stored on ice until further processing.
- If spontaneous expectoration of sputum is not possible, induce sputum as described previously19. Briefly, inhale 200 µg of the β-2-receptor-antagonist salbutamol before starting the sputum induction procedure. Afterwards, inhale a hypertonic (6%) saline solution for 15 min using a nebulizer. Collect the expectorated sputum in a Petri dish.
- Separate the mucus clumps from the saliva into a Petri dish with the help of a pipette tip.
- Weigh the mucus.
NOTE: The average weight of a mucus samples is 0.8 g (varying between 0.1 g and 5 g); 0.1 g are usually sufficient to perform the mentioned procedures. - Add 4 parts (v/w) of 10% Sputolysin (in PBS) to sputum (for example: 4 mL of 10% Sputolysin for each gram of sputum).
CAUTION: Sputolysin is composed of concentrated dithiothreitol in phosphate buffer, therefore handle it with care. - Incubate the mixture at room temperature (RT) on a rocking shaker for 15 min to dissolve the mucus. For safety reasons, place the shaker into a fume hood.
- Quench the reaction by adding the same volume of cold PBS (for example: 1 mL of cold PBS for each mL of 10% Sputolysin).
- Mix by pipetting to obtain a homogenous solution.
- Filter the mixture through a 100 µm nylon cell strainer into a 50 mL tube.
- Repeat the filtration step through a 40 µm nylon cell strainer.
- Centrifuge the solution for 10 min at 300 x g at 4 °C.
- Transfer the supernatant fraction carefully into a fresh tube and store it on ice.
NOTE: The supernatant fractions can be stored at -20 °C or -80 °C until further analysis. - Gently resuspend the cell pellet in 500 µL of cold PBS and place it on ice.
NOTE: The cell fraction must be processed immediately.
2. Neutrophil serine protease activity measurement
NOTE: Here, different methods are introduced to quantify NSPs activity by means of FRET reporters. The choice of the technology is dictated by the specific biomedical question and purpose of the experiment. The probes presented were extensively tested for their specificity against a set of lung relevant enzymes14,15. Although the probes are specific toward their target enzyme, always check the probe specificity on the clinical sample of interest. This can be achieved by incubating the sample with a specific protease inhibitor prior to probe addition, which should abolish any increase in the D/A ratio.
- Soluble NSPs activity quantification via fluorimeter or plate reader assay
NOTE: Protease activity in soluble fractions of the sample can be detected with any instrument capable of fluorescence detection.- Thaw enzymes on ice.
- Until use, keep NE and CG in acidic storage buffer (50 mM sodium acetate, 200 mM NaCl, pH 5.5) to prevent self-cleavage.
- To set up an enzyme standard curve, prepare a 1:2 serial dilution of enzyme (33.9 – 0.271 nM for NE; 42.6 – 0.333 nM for CG) in activation buffer (10 mM Tris-HCl, 500 mM NaCl at pH 7.5). The activation buffer has a neutral pH and therefore enables enzymatic catalysis to occur efficiently.
- To prepare the highest standard concentration (NE concentration: 33.9 nM), dilute 1 µL of NE (33.9 µM) in 999 µL of activation buffer.
- To prepare the second standard (NE concentration: 16.95 nM), mix 200 µL of the first dilution with 200 µL of activation buffer.
- Proceed accordingly to prepare the remaining 1:2 dilutions.
- The last standard, which is the blank, is composed of pure activation buffer. The measurement of technical duplicates or triplicates is recommended. During the standard and sample preparation, try to keep vials on ice.
- In parallel to the standard preparation, dilute sputum samples in activation buffer. Dilute the human samples before assessing their protease activity to remain in the linear range of increase of reporter signal (donor/acceptor ratio). If patient samples were left undiluted, the cleavage would happen too rapidly for a reliable fitting. Since healthy donor sputum contains fewer active proteases compared to samples from CF and COPD patients, different dilutions are generally performed (1:10 for healthy sputum supernatant, 1:20-500 for sputum supernatant of COPD or CF patients).
NOTE: In order to quantitatively measure protease activity in samples where their concentration is unknown, a standard curve with known enzyme concentrations needs to be measured in parallel, ideally on the same plate. The concentration of active enzyme in human sputum is calculated via interpolating the slopes measured in human sputum samples with the ones measured with the standard curves. - Before preparing the samples for measurement, set up the instrument. Set the excitation wavelength for NE FRET probe (NEmo-114) to 354 nm, and set the detection wavelength to 400 nm for the donor and 490 nm for the acceptor. Set the excitation wavelength for CG FRET probe (sSAM15) to 405 nm, and set the emission to 485 (donor) and 580 nm (acceptor).
- Add 40 µL of samples, standard or blank into the wells of a black 96-well half area plate.
- To prepare the master mix containing the reporters (concentration of the reporter in the master mix: 10 µM), dilute the probe stock (1 mM in DMSO) 1:100 in activation buffer. Prepare the needed master mix volume by multiplying 10 µL x the number of required plate wells. To reach the optimal final concentration (2 µM) for fluorescence measurement of NEmo-1 and sSAM reporters, add 10 µL of the master mix to each well (containing 40 µL of either sample, standard or blank) and start the readout. NEmo-1 and sSAM reporters will therefore monitor soluble neutrophil elastase and cathepsin G activity, respectively.
NOTE: If a reagent injector is not available, make sure to start the readout as soon as possible after reporter addition to the samples. - Start the plate reader measurement and record the donor/acceptor ratio increase every 60-90 seconds for at least 20 min or until the increase in the signal reaches a plateau.
- Once data are exported, calculate the donor/acceptor ratio (D/A ratio) by dividing the donor relative fluorescence units (RFU) with the acceptor RFU for each time point and sample.
- Calculate the D/A ratio mean and standard deviation of each sample.
- Determine the slope within the linear growth of the D/A ratio change. The slope is an indicator of the enzyme cleavage rate for a FRET probe. Calculate the concentration of active enzyme in sputum by fitting the linear regression slopes derived from the human samples with the ones calculated from the enzyme standard.
- Membrane-bound NSPs activity quantification via fluorimeter or plate reader assay
- Isolate sputum cells as described above. Resuspend 3 x 104 cells in a volume of 40 µL of PBS. Add the cells to plate reader well.
- Set up the instrument. Set the excitation wavelength for membrane-bound NE FRET probe (NEmo-214) to 405 nm, and set the detection wavelength to 485 nm for the donor and 580 nm for the acceptor. Set the excitation wavelength for membrane-bound CG FRET probe (mSAM15) to 405 nm, and set the emission to 485 (donor) and 580 nm (acceptor).
- To prepare the master mix containing the reporters, dilute the probe stock in activation buffer to a concentration of 10 µM. Prepare the needed master mix volume by multiplying 10 µL X the number of required plate wells. To reach the optimal final concentration (2 µM) for fluorescence measurement of NEmo-2 and mSAM reporters, add 10 µL of the master mix to each well (containing 40 µL of either sample, standard or blank) and start the readout. NEmo-2 and mSAM reporters will therefore monitor membrane-bound neutrophil elastase and cathepsin G activity, respectively.
NOTE: A cellular negative control can be used, for example, cells which do not actively secrete NSPs. For example, the incubation of the reporters with 3 x 104 leukocyte progenitors HL-60 promyelocytic cells represents a valid cleavage negative control. - Record change in donor/acceptor ratio for at least 20 min or until the increase of the signal reaches a plateau. Analyze the data as described above.
- Membrane-bound NSPs activity measurement via fluorescence microscopy
- Determine the number of conditions which need to be analyzed.
NOTE: For each sputum sample measurement, the preparation and analysis of additional positive (PC) and negative (NC) control is recommended. - For each measurement, resuspend 3 x 104 sputum cells in a volume of 50 µL PBS in a 1.5 mL tube.
- As a negative control, incubate sputum cells with a specific inhibitor (Sivelestat, specific NE inhibitor, or cathepsin G inhibitor I, specific CG inhibitor, at a final concentration of 100 µM). Incubate for 10 min at RT.
- As positive control, incubate sputum cells with the appropriate enzyme (NE or CG at 340 nM or 200 nM, respectively) for 10 min at RT.
- Add 50 µL of PBS containing the FRET reporter and a nuclear stain (in a 1:1000 final dilution) to each tube (positive control-treated cells, negative control-treated cells and untreated cells) in order to reach a probe final concentration of 2 µM. Incubate for 10-20 min at RT.
IMPORTANT: The addition of a nuclear stain facilitates fluorescence microscopy imaging as it allows to search for sputum cells of interest without using the FRET probe channels and therefore to avoid reporter bleaching. In addition, the DNA stain permits to segment sputum cells according to the shape of their nucleus. For example, neutrophils can be easily identified by their multilobular nuclei. Also, additional information about the viability of the cells can be retrieved (neutrophils with a more segmented nucleus are more likely to be alive).
NOTE: In addition to the membrane-bound one, the activity of DNA-bound NE or CG in human sputum can be measured in the same way by means of H-NE and H-CG, extracellular DNA-associating FRET probes27. When preparing the master mix, H-NE and H-CG can be added at the concentration of 10 µM and incubated with sputum prior to cytospin and slide preparation. The D/A ratio quantification on extracellular DNA proceeds identically to NEmo-2 and mSAM, with the only difference that extracellular DNA aggregates are segmented instead of single cells27. - Quench the reaction by adding 100 µL of ice-cold PBS, and transfer samples on ice.
- Cytospin the mixture on microscopy slides, air dry, fix with ice cold methanol 10% for 10 min, air dry and mount with an appropriate mounting medium.
NOTE: The microscopy slides can be stored at 4 °C in the dark for up to one month until further analysis. - Acquire microscopy images using a confocal microscope with a PL APO 40x or 63x oil objective. To increase image quality and reduce acquisition time a sequential image acquisition mode is recommended.
- Image the nuclear stain first via 633 nm excitation with the helium-neon-laser line and record its emission between 650 and 715 nm.
- Record the donor (coumarin 343) of the FRET reporter between 470 and 510 nm upon excitation at 458 nm with an argon laser. Acquire the sensitized acceptor (5,6-TAMRA) emission between 570 and 610 nm after sole donor excitation.
- Record the direct acceptor emission in a separate channel between 470 and 510 nm upon acceptor optimal excitation at 561 nm using the diode pumped solid state (DPSS) laser.
- Set the pinhole at the beginning of the experiment and maintain during the course of the imaging session.
NOTE: Due to their small size, the use of a 40x or 63x microscope objective is recommended to visualize neutrophils properly. Usually, images are acquired in several consecutive channels, starting with the longer excitation wavelength to prevent photobleaching of the reporter during acquisition.
- Image at least 100 cells per condition for conclusive statistics. For microscopy images analysis use an appropriate software: segment the cells and calculate the D/A ratio on a pixel-by-pixel basis, afterwards calculate the mean or median of a given region of interest which is selected manually by the user. For representative results see Figure 1.
- Determine the number of conditions which need to be analyzed.
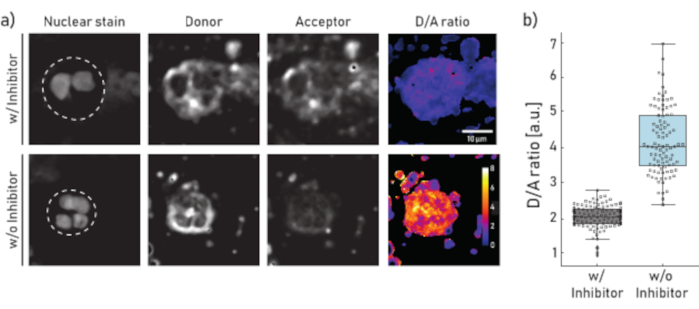
Figure 1: Representative images and quantification of membrane-bound NE activity on neutrophils isolated from CF patient sputum. a) Representative confocal microscopy images of neutrophils pre-incubated (top panel) for 10 min with 100 µM of Sivelestat (w/) or left untreated (w/o) (bottom panel) before reporter NEmo-2 (2 µM) addition. The first column from the left shows the nuclear stain, the second the donor channel, the third the acceptor channel and the last the calculated D/A ratio obtained by dividing donor and acceptor channels on a pixel-by-pixel basis. The borders of the region of interest (single neutrophil) are depicted as dashed line. Scale (10 µm) and calibration bars (D/A ratio) are indicated. b) Box- and dot-plots showing the D/A ratio of sputum neutrophils from a representative CF patient. Cells incubated with inhibitor and untreated cells are shown in grey and blue, respectively. Each dot represents one cell (N: w/ inhibitor = 113 and w/o inhibitor = 96). Please click here to view a larger version of this figure.
- Membrane-bound NSPs activity measurement via flow cytometry
- Resuspend 1 x 106 cells in 100 µL of PBS in a 5 mL FACS polystyrene round-bottom tube and place the tube on ice.
- To gate sputum neutrophils, use the following antibodies: CD14 (1:50), CD16 (1:50), CD45 (1:33) and CD66b (1:50). Prepare sufficient master mix for all samples. Place master mix on ice in the dark.
- Set up the gating strategy as described in Figure 2. The neutrophils are gated as 7AAD–CD45+CD14–CD16+CD66b+ events. The gated events will be then analyzed for their membrane-bound protease activity for their donor (λexc = 405 nm, λem = 450/50 nm) and acceptor (λexc = 405 nm, λem = 585/42 nm) mean fluorescent intensities (MFIs).
- Add 2 µL of FcBlock to each sample, and incubate for 5 min at RT.
- Add the chosen antibodies to each tube, and incubate for 30 min on ice in the dark.
- Wash cells by adding 2 mL of cold PBS, and centrifuge for 5 min at 300 x g and 4 °C. Discard supernatant and resuspend cells in 200 µL of cold PBS.
- Split the 200 µL in two tubes with 100 µL each and add 5 µL of cell viability staining solution in each tube. Place tubes on ice.
- Add an appropriate specific NSP inhibitor (for NE use Sivelestat at 225 µM final concentration, for CG use cathepsin G Inhibitor I at 100 µM) to the negative control (NC) test tube. Incubate the samples at RT for 10 min in the dark.
- Add 100 µL of cold PBS to the sample, filter it through a 40 µm filter in a clean FACS tube to prevent clogging of the instrument.
- Add the reporter (for NE, NEmo-2 at a final concentration of 4 µM, for CG, mSAM at a final concentration of 2 µM) to the NC sample, gently vortex the tube.
- Start acquiring cells incubated with the specific inhibitor to slightly adjust, if necessary, the gates as well as the reporter PMTs' voltages.
- Record at least 1000 neutrophils. Keep the sample at room temperature.
NOTE: Although more events can be recorded, 1000 cells ensure a good compromise between proper statistics and recording time. - Proceed with the following tubes (untreated sputum samples) accordingly.
- To record changes in the D/A ratio due to membrane-bound protease activity, record 1000 neutrophils from each tube every 5 to 10 min.
NOTE: After successful FRET reporter cleavage, the donor channel MFIs intensity will increase over time. The acceptor channel MFIs intensity should decrease or remain constant over time. - Calculate the FRET ratio by dividing the donor by the acceptor channel values for the samples measured on the gated viable single neutrophils.
- Normalize sample-measurements by dividing them with the corresponding 0 min time point (for representative results and analysis see Figure 2).
NOTE: The recording of at least two time points (i.e., 0 and 10 min) is necessary for a dynamic measurement of D/A ratio change. To normalize the activity measurement for each sample, the D/A ratio measured in later time points (e.g., 10 min) is divided by the ratio calculated immediately after probe addition (0 min).
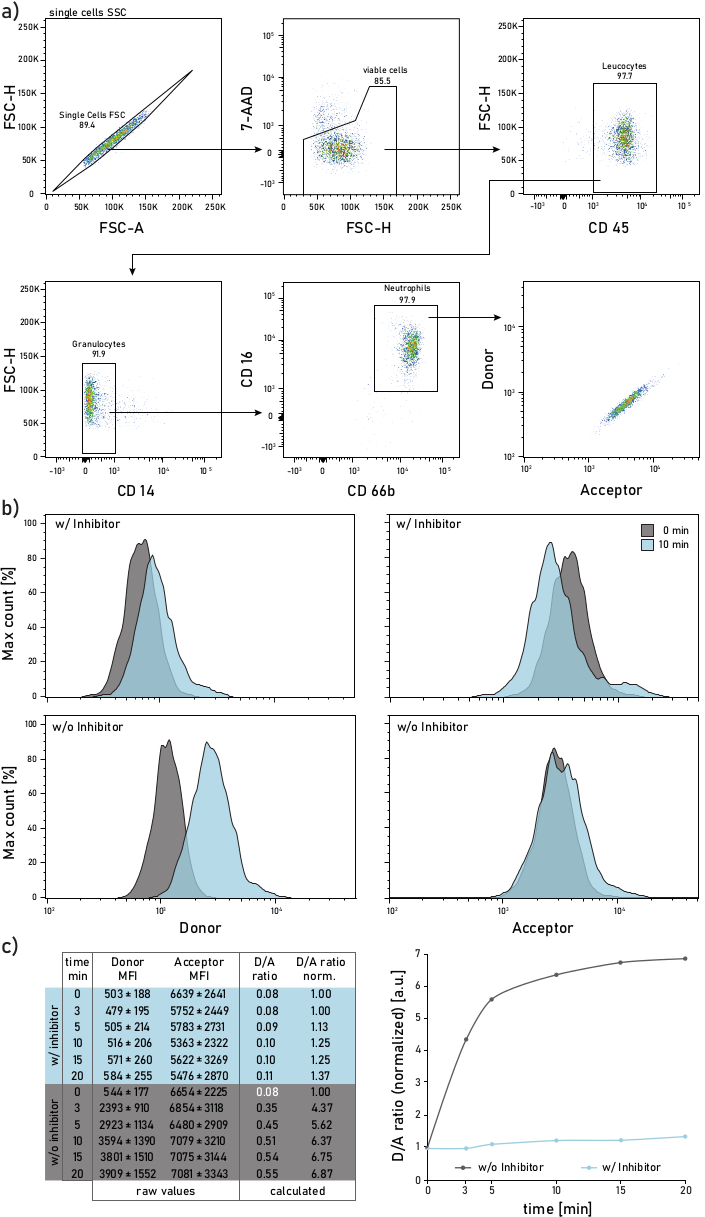
Figure 2: Gating strategy and representative plots of membrane-bound NE activity measured on neutrophils isolated from CF patient sputum. a) To gate sputum neutrophils the following antibodies are used: CD14 (1:50), CD16 (1:50), CD45 (1:33) and CD66b (1:50). The neutrophils are gated as 7-AAD–CD45+CD14–CD16+CD66b+ events. The gated events are analyzed for their donor (λexc= 405 nm, λem= 450/50 nm) and acceptor (λexc= 405 nm, λem= 585/42 nm) mean fluorescence intensities (MFIs). b) Representative histograms of CF sputum neutrophils analyzed for their membrane-bound NE activity. The left column shows the donor signal, the right column shows the acceptor signal. The top row shows mean fluorescence intensities of cells treated with Sivelestat (w/) for 10 min before addition of the reporter. The bottom row shows untreated (w/o) cells whose reporter fluorescence is measured immediately (0 min, grey) and 10 min (blue) after reporter addition. Neutrophils are gated according to the strategy shown in panel a. c) The data table shows a representative dataset consisting of raw MFIs for the donor and acceptor signal on neutrophils measured over several time points (0-3-5-10-15-20 min) as well as the calculated D/A ratio. The D/A ratio can be normalized, i.e., to the 0 min time point (white font). 0 min indicates a recording done as soon as possible after reporter addition to the flow tube with stained sputum cells. MFIs data are shown as mean ± standard deviation for 1000 neutrophils. Please click here to view a larger version of this figure.
