The epithelial SCs reside in a niche called the cervical loop, which is located at the proximal end of the incisor (Figure 3A). Cervical loops are morphologically distinct structures composed of inner and outer enamel epithelium that encase the stellate reticulum, a core of loosely arranged epithelial cells (Figure 3B,C). There are two cervical loops in each incisor (Figure 3A), but only the labial cervical loop contains SCs. The epithelial SCs are localized to the stellate reticulum and the adjacent enamel epithelium at the tip of the cervical loop20,21. They generate progeny characterized by Sfrp5 expression, which differentiate into highly proliferative transit-amplifying cells that produce various cells, including enamel-secreting ameloblasts2,7. Ameloblast differentiation from SCs can be recapitulated in the organ culture system described here2.
In recent years, reporter mice in which a fluorescent protein (such as GFP) is under the control of specific gene regulatory elements have become a widely used tool to identify and isolate cells from various tissues and to follow cell fate and lineage progression in vivo and in vitro22,23. The use of these animal models is beneficial for analyzing the effect of various regulatory molecules, since the intensity and the pattern of fluorescent reporter expression can be used as a readout of endogenous gene activity, or as a reporter of proliferation status (i.e., Fucci reporter mouse model).
The Sox2-GFP transgenic reporter mouse model24, in which the enhanced GFP (EGFP) expression is under the control of a 5.5 kb fragment of the upstream regulatory element of the Sox2 promoter, enables identification and visualization of Sox2-expressing incisor epithelial stem cells (Figure 4A-C). Generation of mice that carry several fluorescent reporters can be useful for identification of more than one cell population. For example, in cervical loops from Sox2-GFP;Fucci-mKO transgenic animals stem cells (GFP+, green, Figure 4D) and non-proliferative cells (Fucci-mKO+, red, Figure 4D) can be identified. In addition, Sox2-GFP expression can be used as a reporter when analyzing the effect of various molecules. Addition of the Wnt/β-catenin signaling activator BIO negatively affects Sox2-expressing stem cells and consequently GFP expression (Figure 4E).

Figure 1: Preparation of the culture chamber. (A) Wash the filter in 1x PBS. (B) Cut the filter into rectangular pieces. (C) Prepare media in the hood and pipet it through the grid. Avoid air bubble formation. (D) Prewarm the pre-prepared culture dish at 37 °C prior to adding tissue explants on a filter placed on the grid. (E) Cartoon representation of the culture chamber. Please click here to view a larger version of this figure.
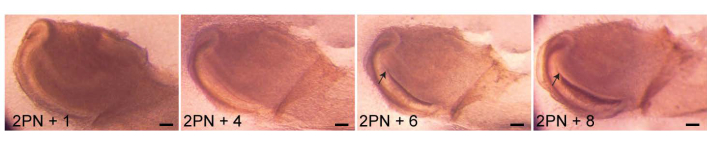
Figure 2: Culturing tissue slices. The proximal end of a 2-day-postnatal (2PN) mouse incisor was dissected and sectioned by vibratome. Slices (150 µm thick) were cultured and hard tissue formation (arrows) was observed after a 6-day culture. Scale 100 µm. Please click here to view a larger version of this figure.
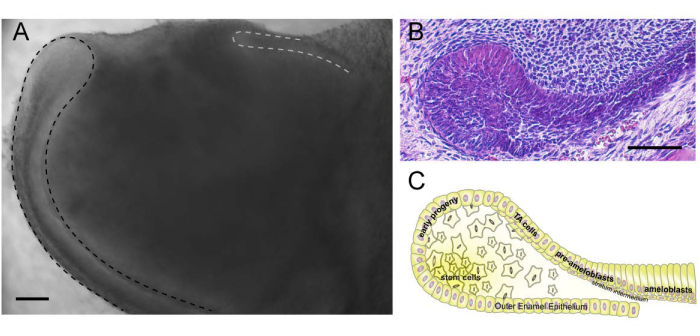
Figure 3: Cervical loop. (A) Proximal end of the incisor-containing labial (outlined by black dashed line) and lingual (outlined by grey dashed line) cervical loops. (B) Histological section of the labial cervical loop, which represents a niche for epithelial stem cells. (C) Cartoon representation of the labial cervical loop and its components. Please click here to view a larger version of this figure.
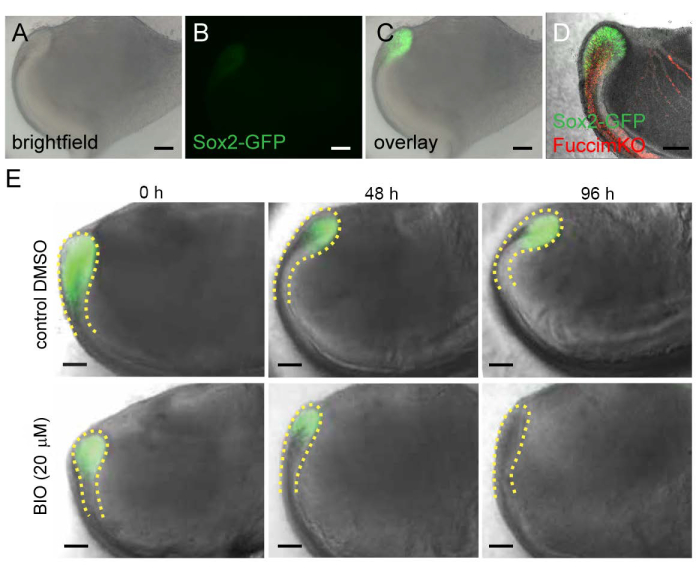
Figure 4: Use of fluorescent reporter mouse models in an organ culture system. (A) Brightfield, (B) fluorescent, and (C) overlay image of the proximal end of the incisor isolated from 2-day postnatal Sox2-GFP mice. (D) Sox2-GFP and Fucci-mKO expression in the proximal end of the incisor isolated from 2-day postnatal Sox2-GFP; Fucci-mKO mice. (E) Effect of BIO on the stem cells expressing Sox2-GFP and the stem cell niche (cervical loop, outlined by yellow dashed line). Scale 100 µm. Please click here to view a larger version of this figure.
