As a proof of concept of the described protocol, the crypts obtained from the Lgr5-eGFP-IRES-CreERT2 mouse line were used in which intestinal stem cells display mosaic GFP expression, which was established by Barker et al., to characterize intestinal stem cells10 initially and allow to map these cells based on their GFP expression. A model is thereby provided to compare ROS levels in a specific cell type population upon different treatments. A ROS inhibitor (NAC) was used, and an inducer (tBHP), known to act on cellular ROS to visualize changes in their levels.
Figures 1A and 1B show representative images of fractions F1 and F4 obtained during the crypts extraction procedure for the intestinal organoid culture. Each fraction must be checked under a microscope or binocular during the extraction procedure to follow crypts detachment and define those fractions enriched in crypts, rather than villi, single cells, or debris. The chosen fractions are then pooled together and passed through a 70 µm cell strainer to remove all the remaining fragments of villi and obtain a preparation with only crypts (Figure 1C). The crypts start to close within a few hours of embedding in BMM, and at D1, round organoids were observed (Figure 1D). After 3-5 days, the organoids will appear with budding structures representing the "newly formed crypts." The organoids are ready for ROS analysis (Figures 1E and 1F).
In the protocol of imaging oxidative stress by confocal microscopy, the slide containing the organoids, incubated with the probe, was imaged with a confocal fluorescence microscope equipped with lasers and filters to detect the Hoechst (ex/em: 361/486), the GFP (ex/em: 488/510) and the fluorogenic probe (ex/em): 644/665) signals. A confocal microscope equipped with 20x air and 63x oil immersion objective allowed the visualization of ROS. In Lgr5-GFP mice, the GFP-positive cells are Lgr5-expressing intestinal stem cells. Supplementary Figure 1 shows representative images obtained with the 20x objective providing an overview of the ROS in several organoids. Figure 3 shows representative images, obtained with the 63x oil objective, of intestinal organoids expressing GFP, non-treated (NT), or pre-incubated or not with the ROS inhibitor NAC, and stimulated or not for 30 min with the ROS inducer tBHP.
In the presence of the inhibitor, the only signal from the dead cells contained in the lumen of the organoid is visible. In the non-treated organoid, the basal ROS levels are shown, proving that stem cells produce higher ROS than differentiated cells (according to the microscope settings, the ROS signal might also be visualized in non-stem cells). GFP-positive cells present a more significant cytoplasmic signal with the inducer in the presence of the fluorogenic probe, demonstrating that ROS levels increase particularly in stem cells after treatment.
Figure 4 shows representative results obtained when analyzing ROS production in intestinal organoids stimulated or not with ROS inhibitor or inducer, using a Flow cytometer equipped with 405 nm, 488 nm, and 630 nm lasers. The gating strategy presented in Figure 4A makes it possible to evaluate ROS production at the level of the whole organoids cell population, defining intact and living cells based on physical parameters and DAPI exclusion (SSC-A vs. FSC-A and DAPI vs. FSC-A) and FSC-H vs. FSC-A) or only in the intestinal stem cells, further gated on cells with GFP high signal. Figure 4B shows the ROS levels in the total population upon collection of 50,000 events. Basal ROS levels in the non-treated (NT) cells decrease after stimulation with the inhibitor (NAC), and on the contrary, increase after challenge with the inducer (tBHP). Cells pre-treated with the inhibitor and then stimulated with the inducer present a lower level than those stimulated with the inducer alone. The results were then analyzed using appropriate software, obtaining the median fluorescent intensity (MFI). The obtained values are presented as a ratio over the non-treated cells, as shown in the graph presented on the right of Figure 4B. Figure 4C shows the same parameters described in Figure 4B in the stem cells, gated as GFP positive cells, showing a 3.5-fold decrease in ROS level upon NAC treatment and 4-fold increase upon tBHP treatment over non-stimulated cells. This result demonstrates that following this protocol, it is possible to quantify differences in ROS levels at the level of the whole cell population or in GFP positive stem cells upon their treatment of the organoids with specific compounds.
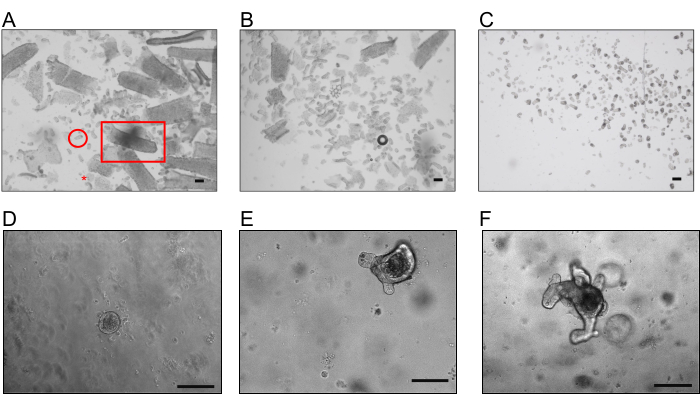
Figure 1: Representative images of crypts and organoids. (A) Example of fraction F1 obtained after the first incubation with EDTA, enriched in villi (square), with some debris (star) and crypts (circle). (B) Example of fraction F4 enriched in crypts. (C) Suspension presenting only isolated crypts obtained after the filtration with a 70 µm cell strainer (scale bar, 200 µm). (D, E, and F). Typical organoids were obtained after 1, 3, and 5 days respectively, after embedding the crypts in BMM (scale bar, 100 µm). Please click here to view a larger version of this figure.
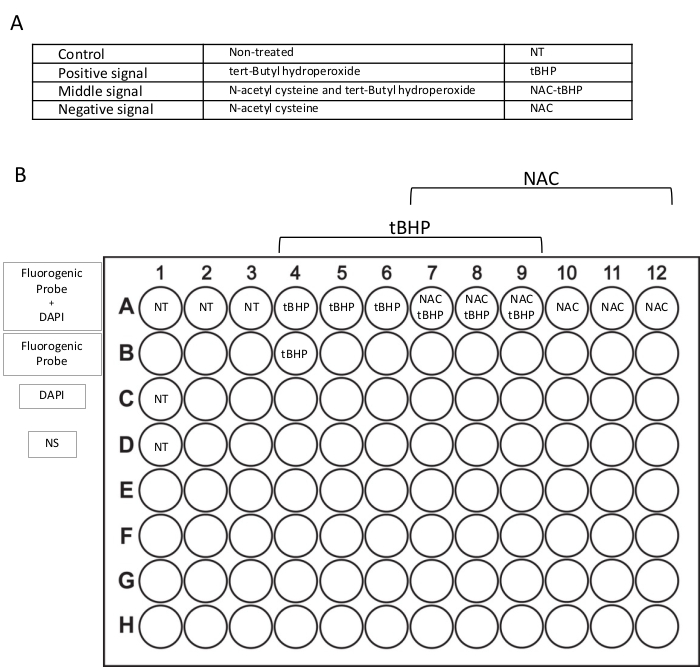
Figure 2: Outline of the experimental plan. (A) Conditions used in this protocol included in each experiment: non-treated wells (NT), inducer-treated wells (tert-Butyl hydroperoxide – tBHP), inhibitor-treated wells (N-acetyl cysteine – NAC), and inhibitor- and inducer-treated wells (NAC-tBHP). (B) Plate format for the flow cytometry assay. Each condition is plated in triplicate (line A). Lines B, C, and D include wells for flow cytometer setting with only the fluorogenic probe, only DAPI, or non-stained (NS) samples. Please click here to view a larger version of this figure.
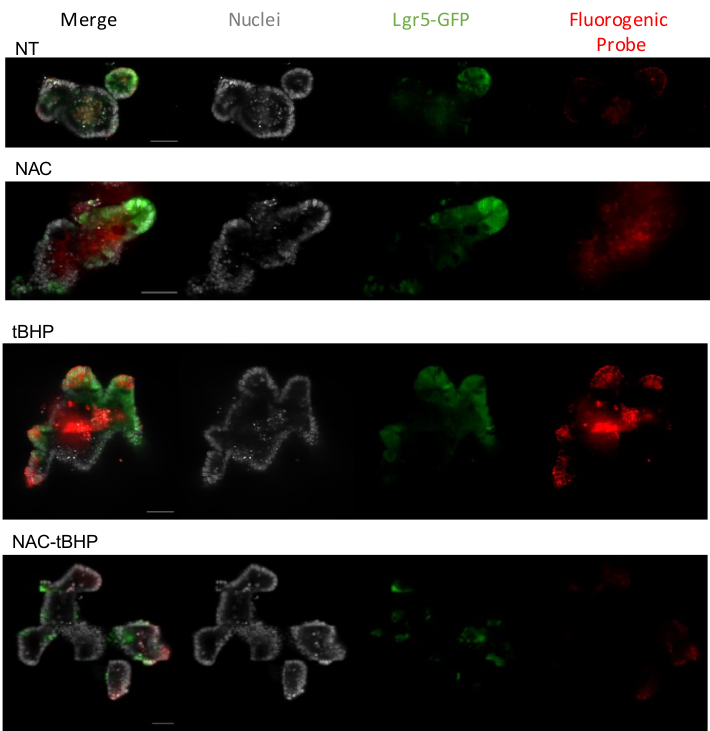
Figure 3: Representative confocal images of ROS staining in organoids. Stitched images were obtained with a confocal microscope equipped with a high-speed EMCCD Camera, 63x/1.4 oil objective, and slit 35 µm, using the lasers 405, 488, 640, and filters 460/50, 535/50, 700/75 to acquire Hoechst, GFP, and the fluorogenic probe respectively. Confocal optical sections of organoids non-treated (NT), treated with the ROS-inhibitor (NAC), with the ROS-inducer (tBHP), or pre-treated with the ROS inhibitor and then stimulated with the ROS-inducer (NAC-tBHP). In grey, nuclei stained with Hoechst; in green, Lgr5-GFP cells; in red, the fluorogenic probe (scale bar, 50 µm). Please click here to view a larger version of this figure.
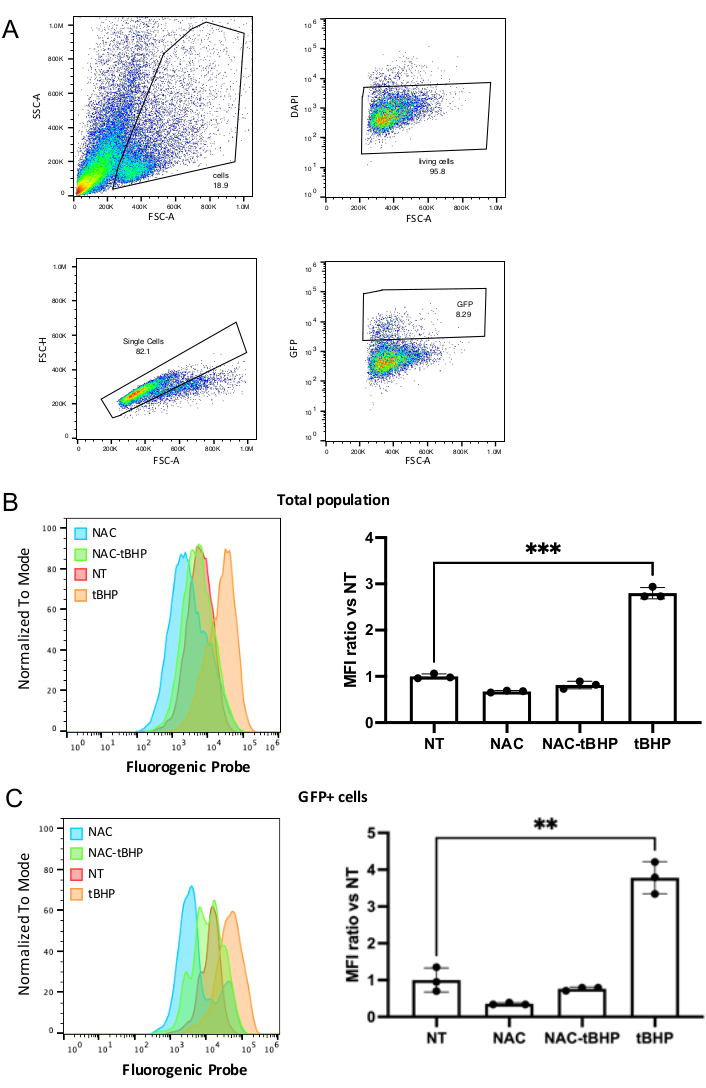
Figure 4: Representative flow cytometry analysis of ROS in cells derived from organoids. (A) Schematic representation of the gating strategy used in flow cytometry analysis: gating for cell shape (exclusion of dead cells and debris accumulated in the organoids lumen), gating for living cells (cells not incorporating DAPI-laser 405), gating for single cells (doublet discrimination), and stem cells (GFP positive cells-laser 488) (FSC: forward scatter, SSC: side scatter). The ROS signal has been acquired using the 630 laser. (B) On the left, histograms were obtained with an appropriate software showing the intensity ROS signals for the total living population (after gating around 10,000 events per condition) in the different samples NT: non-treated; NAC: inhibitor-treated; tBHP inducer-treated; NAC-tBHP: inhibitor- and inducer-treated. On the right, a typical example of the calculated ratio for MFI values over the NT samples obtained during an experiment starting from 3 samples per condition (mean ± SD) (*** P = 0.0003). (C) Same as in B for the GFP positive population (1,000 events per condition) (* P = 0.02). Please click here to view a larger version of this figure.
Supplementary Figure 1: Representative confocal images of ROS staining in organoids. Stitched images were obtained with a confocal microscope equipped with a high-speed EMCCD Camera, 20x objective, and slit 35 µm, using the lasers 405, 488, 640, and filters 460/50, 535/50, 700/75 to acquire Hoechst, GFP, and fluorogenic probe respectively. Confocal optical sections of organoids non-treated (NT), treated with the ROS-inhibitor (NAC), with the ROS-inducer (tBHP), or pre-treated with the ROS inhibitor and then stimulated with the ROS-inducer (NAC-tBHP). In grey, nuclei stained with Hoechst; in green, Lgr5-GFP cells; in red, fluorogenic probe (scale bar, 100 µm). Please click here to download this File.
