All mouse protocols were approved by the Animal Care and Use Committee of the National Eye Institute (NEI ASP# 650). Mice were housed in 12 h light-dark conditions and cared for by following the recommendations of the Guide for the Care and Use of Laboratory Animals, the Institute of Laboratory Animal Resources, and the Public Health Service Policy on Humane Care and Use of Laboratory Animals.
1. Hydrating sensor cartridge and preparation of the assay medium
- The day before the experiment, add 1 mL of the calibration medium to each well of the utility plate. Place the Hydro-Booster cover on the top and lower the sensor cartridge through the opening on the cover. Check to ensure that the sensor is submerged in the calibration medium. Incubate the sensor cartridge overnight in a CO2-free incubator at 37 ˚C to activate the fluorophores.
NOTE: To prevent evaporation, the incubator is humidified by keeping a tray of water inside, and the sensor cartridge cassette is wrapped with clear plastic wrap. - Prepare the assay medium by reconstituting the Seahorse DMEM medium with the addition of glucose, pyruvate, and glutamine to the desired concentrations. In the assays reported in this article, the final concentration of substrates in the assay medium are: 6 mM of glucose, 0.12 mM of pyruvate, and 0.5 mM of glutamine. For each assay plate, 40 mL of the assay medium is prepared fresh on the day of the experiment.
- Set up the assay program in the analyzer following the manufacturer's instruction26. In the assay demonstrated here, the protocol is set as follows: 5 cycles of measurements for baseline, then inject port A, followed by 4 cycles of measurements, then inject port B and followed by 4 cycles of measurements. Each cycle is composed of mix (3 min), wait (2 min) and measure (3 min).
2. Coating mesh inserts of islet capture microplate
- Prepare the coating mix by combining 20 µL of the cell attachment medium (e.g., Cell-Tak) with 171 µL of 0.1 M sodium bicarbonate and 9 µL of 1 M NaOH.
- Open the lid of the cassette containing mesh inserts. Pipette 8 µL of the coating mix to each mesh inserts. Use a pipette tip to gently smear/spread the droplet around to distribute the coating mix equally throughout the mesh insert.
- Close the cassette and allow the mesh inserts to incubate at room temperature for at least 25 min for adsorption.
- Wash the mesh insert by pipetting 4 mL of the assay medium directly onto the mesh inserts. Gently shake the cassette to ensure all mesh inserts are washed with the assay medium.
- Keep the mesh insert aside. It is ready to use.
3. Preparing injection compounds
- Take out stock aliquots of Bam15 (10 mM), Rotenone (10 mM), Antimycin A (10 mM) and 2-DG (500 mM) from -80 °C freezer and thaw at room temperature.
NOTE: The 2-DG stock is ready to use. The other drugs need to be diluted to working stock. - Warm up 10 mL of the assay medium in a 37 °C water bath.
- Dilute 10 mM Bam15 stock to 50 µM working stock using a two-step dilution procedure: mix 20 µL of 10 mM stock with 20 µL of DMSO to get 5 mM intermediate stock. Then mix 10 µL of the above 5 mM intermediate stock with 990 µL of pre-warmed assay medium to get the final 50 µM working stock.
- Dilute and combine 10 mM Rotenone and 10 mM Antimycin A stock to 10 µM Rotenone/Antimycin A (Rot/AA) working stock by two steps of dilutions: mix 10 µL each of 10 mM Rotenone and 10 mM Antimycin A stock with 80 µL of DMSO to get 1 mM Rot/AA intermediate stock. Then mix 10 µL of the above 1 mM intermediate stock with 990 µL of pre-warmed assay medium to get the final 10 µM Rot/AA working stock.
- Freshly prepare the above-mentioned working stocks of injection compounds on the day of the experiment and set them aside at room temperature until loading into injection ports of the sensor cartridge.
4. Retinal dissection and retinal punch preparation
- Euthanize a mouse by CO2 asphyxiation following AVMA Guidelines on Euthanasia27.
NOTE: Do not leave the animal in a CO2 chamber longer than the time needed for euthanasia. - Enucleate eyes and place into ice-old 1x PBS buffer in a Petri-dish and then place it under a dissection microscope.
- Carefully remove, by cutting with microscissors, the extra rectus muscles attached outside the eyeball and cut off the optic nerve.
- Use a 30 G needle to punch a hole at the edge of the cornea (limbus); this serves as the insertion site for the microscissors. Then, use a fine dissection microscissors to make a circular cut along the edge of the cornea, separating it from the posterior eye cup.
- Use sharp dissection forceps to remove the cornea, lens, and the vitreous humor away from the eye cup.
- Use fine dissection microscissors to make several small cuts on the scleral layer at the rim of eye cup. Avoid cutting the retina layer. Use two sharp dissection forceps to hold on to the scleral tissue at each side of the cut and very carefully pull on the scleral layer to remove it from the neural retina. Repeat this around the eye cup until all sclera is removed and an intact retinal cup is obtained.
- Use dissection microscissors and make radial cuts on the retinal cup to flatten it and generate several distinct sections.
NOTE: Depending on the person's dissection skills and experience in handling fresh retinal tissue, the retinal cup can be cut to generate 3 to 5 distinct sections. - Use 1 mm diameter biopsy puncher to cut one retinal disk from each section of the flattened retinal cup.
NOTE: Care should be taken to get the retinal disks punched at equal distance from the optic nerve head. - Use forceps to transfer the pre-coated mesh inserts into the dissecting petri-dish. With the help of two superfine eyelash brushes, place the retinal punch disk onto the mesh insert. The retinal punch disk is placed at the center of the mesh insert with ganglion cell layer side down touching the mesh and photoreceptor layer facing up.
NOTE: Frequently, some RPE cells remain attached to the photoreceptors, and the pigmentation of these cells can be used as an indicator of the retinal punch disk orientation.
5. Loading the sensor cartridge injection ports and calibration
- Take the hydrated sensor cartridge plate cassette out of the 37 °C incubator. Remove the Hydro-Booster cover and place the sensor cartridge back on the utility plate.
- Load the desired volume of injection compound working stock solutions into appropriate ports. Hold the pipette tip at 45 ° angle. Insert the pipette tip halfway into an injection port with the bevel of the tip against the opposite wall of the injection port and gently load the compound into each port. Avoid introducing air bubbles.
- Refer to the instrument user guide for the volume of the compound loaded in each injection port for a specific assay. In the experiments presented in this paper, 68 µL of 50 µM Bam15 working stock (for mitochondrial stress assay) or 68 µL of 10 µM Rot/AA working stock (for glycolytic rate assay) is loaded into port A; 75 µL of 10 µM Rot/AA working stock (for mitochondrial stress assay) or 75 µL of 500 mM 2-DG working stock (for glycolytic rate assay) is loaded into port B.
- Load injection ports of all wells of the plate including background correction wells and blank wells to ensure proper injection. Load the respective compound solution in each port for the background correction wells. Assay medium can be substituted, instead of the compound solution, in each of the ports of the bank wells.
- Place the loaded sensor cartridge plate, with lid off, into the analyzer machine to start calibration prior to the assay run. After the calibration is over, the program will automatically pause, waiting for the replacement of the utility plate with the islet capture plate containing retinal punches.
6. Loading the islet capture plate and start assay run
- Add 607 µL of the assay medium to each well of the islet capture plate
- Use forceps to grab the rim of the mesh insert containing retinal punch disks on top and take it out from the Petri-dish. Lightly tap the bottom of the mesh insert on an absorbing wipe tissue to remove extra liquid and put it into the well of the islet capture plate. Repeat this step until all mesh inserts with retinal punches are placed into the islet capture plate. Fill background correction wells and blank wells with empty mesh inserts.
- Use two Graefe forceps to carefully and gently press the rim of each mesh insert and make sure that these are securely inserted at the bottom of the islet capture plate.
- Place the loaded islet capture plate into a 37 °C incubator for 5 min to warm up.
- Eject the utility plate after the calibration is complete and replace it with an and replace it with the islet capture plate, with lid off, containing retinal punches.
- Resume the assay run.
7. Run termination and data storage
- After the run is complete, eject the sensor cartridge and islet capture plate containing retinal punches. The data is automatically saved as .asyr file.
- Use the associated data analysis software to view and analyze the data following the manufacturer's user guide26.
- Use the Export function to export .xslx file of the data, which can be viewed and analyzed using spreadsheet software.
8. Saving the retinal punch sample
- After the assay, take out the plate from the machine, remove the sensor cartridge and gently remove the assay medium from each well using a pipette.
- Apply the cover back on and seal the sides of the plate with the parafilm strip.
- Store at -80 °C.
- For normalization, quantify the total DNA or protein content of the punch in each well.
9. Data analysis
- Mitochondrial stress assay
NOTE: The measured OCR value (totalOCR) represents total oxygen consumption by the tissue. After Bam15 (uncoupler) injection, OCR increases from the basal level (totalOCRbasal) to the maximum level (totalOCRmax) and goes down following the Rot/AA injection. The residual OCR value after Rot/AA injection (totalOCRRot/AA) represents non-mitochondrial oxygen consumption.- Calculate mitochondria-related oxygen consumption as:
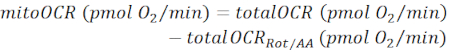 (Eq. 1)28
(Eq. 1)28 - Calculate the mitochondrial reserve capacity (MRC) as:
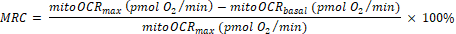 (Eq. 2)29
(Eq. 2)29
NOTE: The last reading among the 5 measurements before Bam15 injection is taken as the "basal" value (for totalOCRbasal and mitoOCRbasal). The highest reading among the 4 measurements following Bam15 injection is used as "max" value (for totalOCRmax and mitoOCRmax). The lowest reading among the 4 measurements following the Rot/AA injection is used as totalOCRRot/AA.
- Calculate mitochondria-related oxygen consumption as:
- Glycolytic rate assay
NOTE: The measured ECAR value (totalECAR) represents the total acidification of the medium by the tissue's metabolic activity. In general, acidification of the extracellular micro-environment results mainly by extrusion of the glycolytic product, lactate. Catabolism of substrates in mitochondrial TCA cycle results in the production of CO2, which also acidifies the extracellular medium through hydration to bicarbonate.- Substract mitochondrial contributed medium acidification (mitoECAR) from totalECAR to obtain the glycoECAR.
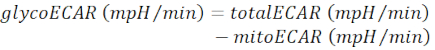 (Eq. 3)28
(Eq. 3)28
NOTE: Mitochondrial respiration and TCA cycle are strongly coupled processes. Production of CO2 from mitochondria is a function of the rate of OXPHOS, which is measurable by mitoOCR. - Calculate the mitoECAR as:
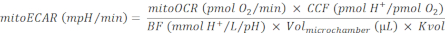 (Eq. 4)28
(Eq. 4)28
where, the CCF (CO2 Contribution Factor) is an empirically calculated ratio value, representing the amount of H+ contribution from CO2-mediated acidification vs each O2 consumption from OXPHOS. CCF for this system is pre-determined to be 0.6028. Accurate measurement of medium acidification is determined by the buffer capacity of the medium, the sensitivity of instrument pH sensor, and the effective measurement chamber capacity. Here, the BF (Buffer Factor) is a parameter of the in situ experimental buffer capacity, representing the amount of H+ or OH– added to the effective measurement chamber to change the pH level by 1 unit. When customized assay medium is used, the BF can be determined by titrating known amounts of acid into the assay medium following the Buffer Factor protocol30. The Seahorse DMEM medium pH 7.4 used in this protocol has a pre-determined BF of 2.60 mmol H+/L/pH. The islet capture plate used in this protocol has a Volmicrochamber = 16.6 µL31. The volume scaling factor, Kvol, is an empirically determined constant. Kvol value is not available for the islet capture plate but can be calculated from the value of the microplate28, accounting for the volume difference in their microchambers, to be 0.41.
NOTE: Injection of the Rot/AA shuts down mitochondrial respiration and forces the tissue to switch to glycolysis for ATP production, leading to higher lactate extrusion and an increase in ECAR measurement. Glycolysis is ceased with 2-DG injection, and the residual ECAR measurement reveals non-glycolytic and non-mitochondrial acidification of medium. - Calculate the glycolytic reserve capacity (GRC) as:
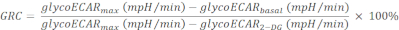 (Eq. 5)32
(Eq. 5)32
where, the last reading among the 5 measurements before Rot/AA injection is taken as the "basal" value (glycoECARbasal). The highest reading among the 4 measurements following Rot/AA injection is used as "max" value (glycoECARmax). The lowest reading among the 4 measurements following 2-DG injection is used as glycoECAR2-DG.
- Substract mitochondrial contributed medium acidification (mitoECAR) from totalECAR to obtain the glycoECAR.
- Normalization
NOTE: Normalization is essential when comparing the readings from retinal tissues of different age groups or between wild-type and pathological/degenerative samples, which might differ in cell numbers.- Use commerically available kits to assess the DNA content in each retinal punch disk33,34.
- Alternatively, use radioimmunoprecipitation assay buffer (RIPA buffer) to extract total protein from the retinal punch and use the protein content for normalization.
NOTE: The surface area of an adult mouse retina has been previously determined to be around 20 mm2, and each retina contains ~6.5 million cells35. Hence, each 1 mm diameter retinal punch is ~1/25 of a single retina and contains ~260K cells. One can refer to these numbers when comparing the data from a retinal punch to those from other tissue samples or cultured cells,
The data reported here are representative mitochondrial stress assay showing OCR trace (Figure 1) and glycolytic rate assay showing OCR trace and ECAR trace (Figure 2), which were performed using freshly dissected 1 mm retinal punch disks from 4 months old transgenic Nrl-L-EGFP mice36 (C57B/L6 background). These mice express GFP specifically in rod photoreceptors without altering normal retinal development, histology, and physiology and have been widely used as wild-type controls in retinal research. Two Nrl-L-GFP littermate mice were used were used in the assays presented here. GFP expressed in the Nrl-L-GFP mice does not interfere with the measurements of OCR and ECAR in this protocol. Five retinal punches were taken from each retina. Ten of the retinal punches were used for mitochondrial stress assay and the other 10 were used for glycolytic rate assay. Seahorse XF DMEM medium, pH 7.4 (constituted with 6 mM glucose, 0.12 mM pyruvate, and 0.5 mM glutamine) and Seahorse XFe24 Islet Capture plates were used in the experiments. The representative data presented here were obtained using the same 1 mm diameter puncher but was not normalized with respect to the DNA/protein content.
In mitochondrial stress assay, the uncoupler Bam1537 was injected after establishing the OCR baseline, leading to enhanced OCR to the maximal level. Rotenone and Antimycin A were injected to inhibit mitochondria respiration at complex I and complex III, respectively, resulting in OCR to drop to the minimal level (Figure 1). The difference between the maximal level of OCR and the last measurement of the basal OCR level reflects mitochondrial reserve capacity (MRC). The MRC is calculated to be 19.2%±3.4% using Eq. 2, consistent with previously measured MRC values in retinas of ~3 months old Nrl-L-EGFP mice using the previous generation Seahorse XF24 analyzer22,38.
In the glycolytic rate assay, Rotenone and Antimycin A were injected after establishing the baseline for the total ECAR. With the production of ATP from OXPHOS halted, the tissue is forced to rely on glycolysis for energy, and an increase in the extracellular release of lactate drives ECAR to the maximal level. Glycolysis is ceased by injection of 2-DG, which competes with glucose for hexokinase binding, causing ECAR to drop to the minimal level (Figure 2). Mitochondria contributed ECAR (mitoECAR) can be calculated from the mitoOCR value (Eq. 4). Glycolysis contributed ECAR glycoECAR is calculated and plotted by subtracting mitoECAR from totalECAR. The difference between maximal level of glycoECAR and the last measurement of glycoECAR basal level reflects the glycolysis reserve capacity (GRC). Here, the GRC is calculated to be 35.7% ± 3.4% using Eq. 5.
As a highly glycolytic tissue, lactate production from the retina accounts for a major source of extracellular acidification, as revealed by the small difference of glycoECAR from the totalECAR. Interestingly, ECAR measurement does not plateau immediately following the Rot/AA injection but drops after the second measurement. The retinal punch disk is an intact ex vivo system composed of different cell types, including the Müller glia cells, which are known to receive lactate (glycolysis end product) released from the photoreceptors6. Hence, a drop in ECAR measurement following the Rot/AA injection is likely explained by increased removal of lactate from the intercellular space, slowing down/preventing its release into the medium.
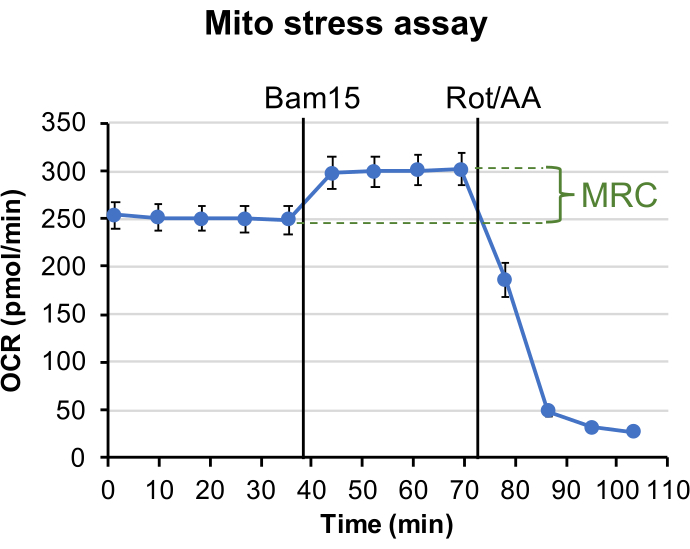
Figure 1: Mitochondrial stress assay. The plotted graph shows OCR trace from 1 mm retinal punch disks in Seahorse XF DMEM buffer, supplemented with 6 mM of glucose, 0.12 mM of pyruvate and 0.5 mM of glutamine. Each data point represents the average of measurements from 10 wells. Error bar = standard error. MRC is calculated to be 19.2%±3.4%. Please click here to view a larger version of this figure.
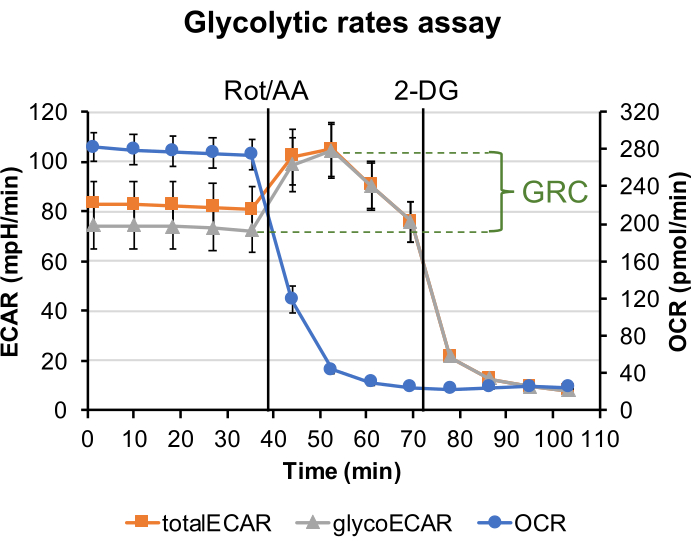
Figure 2: Glycolytic rate assay. The plotted graph shows the measured OCR trace, ECAR trace (totalECAR), and the calculated glycolysis contributed ECAR (glycoECAR) from 1 mm retinal punch disks in Seahorse XF DMEM buffer supplemented with 6 mM of glucose, 0.12 mM of pyruvate, and 0.5 mM of glutamine. Each data point represents the average of measurements from 10 wells. Error bar = standard error. GRC is calculated to be 35.7%±3.4% Please click here to view a larger version of this figure.
