After multiple fluorescent staining, the corresponding labeling was orderly demonstrated on the 80 µm thick sections of the rat medial gastrocnemius muscle with NF200-positive nerve fibers, VAChT-positive pre-synaptic terminals, α-BTX-positive post-synaptic AchRs, S100-positive PSCs, phalloidin-positive muscular fibers, and DAPI-labeled cellular nuclei (Figure 3 and Figure 4).
It was shown that NF200-positive nerve fibers ran in bundle and gave out branches to the VAChT-positive pre-synaptic terminals that formed a mirror relationship with the α-BTX-positive post- synaptic AchRs (Figure 3). In addition, S100-positive PSCs were detected around the NF200-positive nerve fibers close to the pre-synaptic terminals (Figure 4).
By taking advantage of the image-reconstruction, the spatial correlation of the nerve fibers, pre-synaptic terminals, PSCs, and post-synaptic AchRs was further exhibited in a three-dimensional pattern showing the detailed morphological characteristics of NMJ from different perspectives (Figure 3C,C1 and Figure 4C,C1).
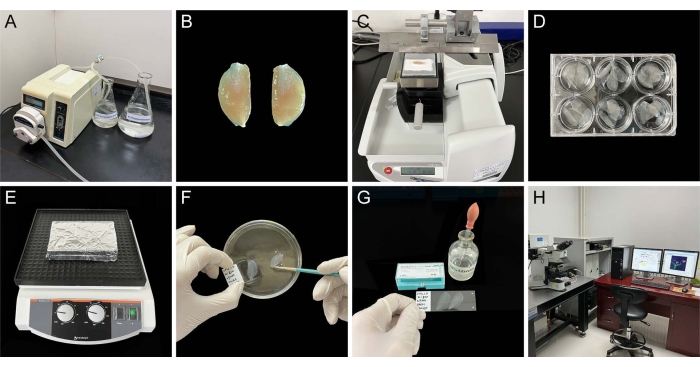
Figure 1. Images of the various protocol steps. (A) Perfuse the rat in the hood. (B) Dissect out the rat medial gastrocnemius muscle. (C) Cut the muscular tissue on a freezing stage of sliding microtome system. (D) Collect tissue sections orderly in a six-well dish. (E) Fluorescent staining on the shaker. (F) Mount the sections on microscope slides. (G) Apply coverslips to the sections with 50% glycerin. (H) Observe the samples with a confocal imaging system. Please click here to view a larger version of this figure.
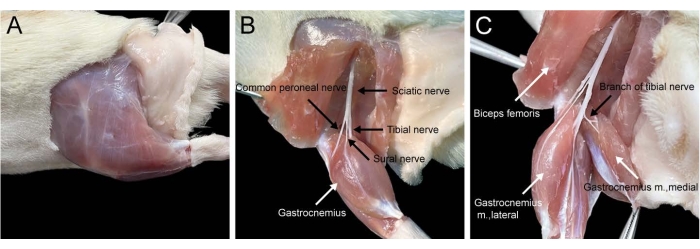
Figure 2. Procedure of regional anatomy for dissecting out the rat medial gastrocnemius muscle. (A) Outside view of muscles in the rat hind limb. (B) Inside view of the medial gastrocnemius muscle and its innervation. (C) Dissecting out the rat medial gastrocnemius muscle. Please click here to view a larger version of this figure.
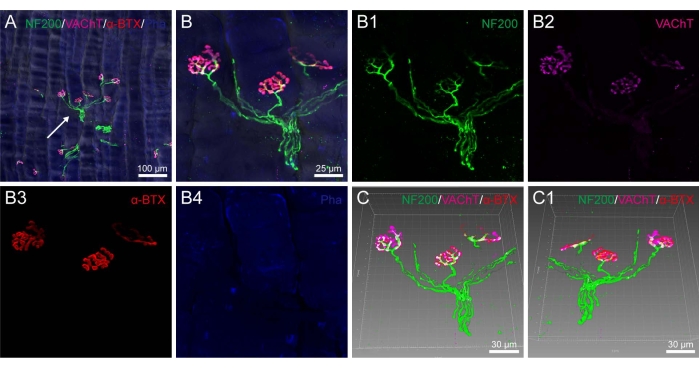
Figure 3. Spatial correlation of the nerve fibers, pre-synaptic terminals, post-synaptic nicotinic acetylcholine receptors, and muscular fibers on the rat medial gastrocnemius muscle. (A) Representative image from the rat medial gastrocnemius muscle showing the multiple fluorescent staining with neurofilament 200 (NF200), vesicular acetylcholine transporter (VAChT), alpha-bungarotoxin (α-BTX), and phalloidin (Pha). (B) The magnified image from panel A (arrowhead) showing the various labeling in detail. B1-B4: panel B shown separately with NF200 (B1), VAChT (B2), α-BTX (B3), and Pha (B4). (C) The adjusted images from panel B with the frame showing the front view of the 3D pattern in (C) and the back view in (C1). Same scale bar for B1-B4 as in B. Please click here to view a larger version of this figure.
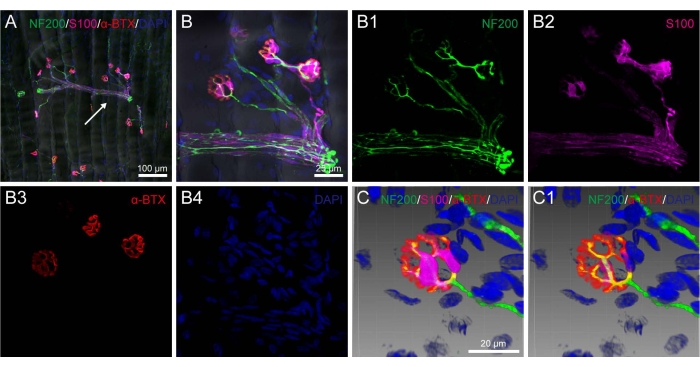
Figure 4. Spatial correlation of the nerve fibers, peri-synaptic Schwann cells, post-synaptic nicotinic acetylcholine receptors, and cellular nuclei on the rat medial gastrocnemius muscle. (A) Representative image from the rat medial gastrocnemius muscle showing the multiple fluorescent staining with neurofilament 200 (NF200), Schwann cell marker (S100), alpha-bungarotoxin (α-BTX), and cellular nucleus marker 4',6-diamidino-2-phenylindole dihydrochloride (DAPI). (B) The magnified image from panel A (arrowhead) showing the various labeling in detail. B1-B4: show panel B separately with NF200 (B1), S100 (B2), α-BTX (B3), and DAPI (B4). (C) The adjusted images from panel B with the frame in a 3D pattern in (C) and the view without S100 labeling in (C1). Same scale bar for B1-B4 as in B. Please click here to view a larger version of this figure.
