NOTE: This protocol has two main steps: a wet lab step, involving cell culture and live confocal microscopy to obtain images of live mitochondria (Figure 1) and an in silico step to analyze obtained images (Figure 2). For automated data analysis of 3D live imaged mitochondria, the MATLAB application Mitometer was used as provided by Lefebvre et al.9. The Routine optimization is written in MATLAB. The software, updated versions and processing ImageJ Macros are freely available online through GitHub, at https://github.com/JoseJoaoMV/Routine_Optimization_Mitometer_APP_MATLAB.
1. Live microscopy
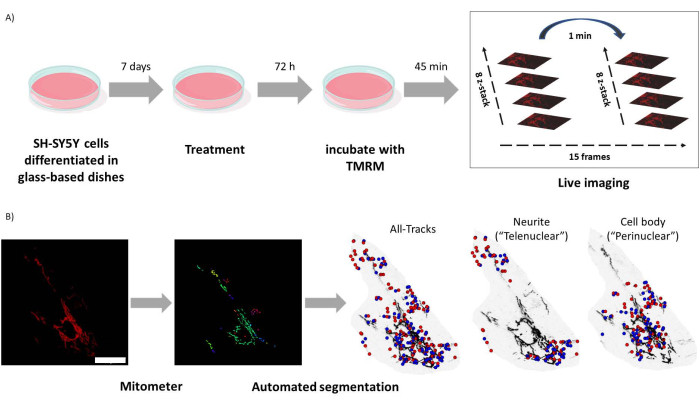
Figure 1: Experimental protocol. SH-SY5Y cells were differentiated and treated with retinoids. (A) TMRM was used to live image healthy mitochondria in treated cells using a confocal microscope, capturing a time lapse z-stack of five visual fields. (B) Mitometer application MATLAB automatically segments and analyzes mitochondria images. In addition to analyzing, this software automatically discriminates mitochondria according to nuclear proximity. Blue dots are mitochondrial initial positions; red dots are final positions. Scale bar = 30 µm. Abbreviation: TMRM = tetramethylrhodamine, methyl ester. Please click here to view a larger version of this figure.
- Cell culture
- Incubate SH-SY5Y cells at 37 °C in a humidified atmosphere of 5% CO2 and 95% air, cultured in equal parts of MEM (Minimal Essential Medium) and F12 (Ham's F12 Nutrient Mix) Medium, supplemented with 10% fetal bovine serum (FBS).
- Plate SH-SY5Y cells in glass-bottom cell dishes at a density of 15 × 104 cells/mL.
- Differentiate cells with 5 days of treatment with 10 µM all-trans retinoic acid in 1% FBS-containing culture medium, followed by 2 days of treatment with 10 ƞg/mL brain-derived neurotrophic factor (BDNF).
- Cell treatment
- Wash cells with sterile phosphate-buffered saline (PBS) and treat for 72 h with 10-7 M RAR-isoform agonists, in equal parts of MEM (Minimal Essential Medium) and F12 (Ham's F12 Nutrient Mix) Medium, supplemented with 1% FBS.
NOTE: The RARα agonist used was AM580; RARβ agonist used was CD2314; Ch55 was used as agonist as RARα and β co-agonist; atRA was used as a positive control; BMS493 was used as a RAR pan-antagonist.
- Wash cells with sterile phosphate-buffered saline (PBS) and treat for 72 h with 10-7 M RAR-isoform agonists, in equal parts of MEM (Minimal Essential Medium) and F12 (Ham's F12 Nutrient Mix) Medium, supplemented with 1% FBS.
- Live confocal imaging
- Replace culture medium with fresh 1% FBS-containing culture medium with 20 nM tetramethylrhodamine, methyl ester (TMRM) for 45 min.
NOTE: TMRM is a cell-permeant fluorescent dye, sequestered by active mitochondria, and this incubation period allows TMRM to reach an equilibrium and be taken up by polarized mitochondria11. Imaging should be started before equilibrium is established as TMRM signal intensity could artificially increase during imaging. - Place the cells in an incubator attached to a laser-scanning confocal microscope at 37 °C.
- Capture images using a 63x oil-immersion apochromat objective, with an image size of 512 x 512 pixels obtained with a pinhole aperture of 1 airy unit, capturing a time series of 15 frames from five different visual fields in each cell plate, and a z-stack of 8 equidistant z-planes. The resulting .lsm file is a time-, position-, and z-stack.
NOTE: Settings for gain, contrast, and brightness must be initially optimized using the minimum laser power necessary to use the entire dynamic range of the detector and kept constant throughout the study, ensuring that all imaging is performed under the same conditions. Up to nine different positions can be recorded in this hardware-software combination and the microscope cycles between imaging positions automatically.
- Replace culture medium with fresh 1% FBS-containing culture medium with 20 nM tetramethylrhodamine, methyl ester (TMRM) for 45 min.
2. Image analysis
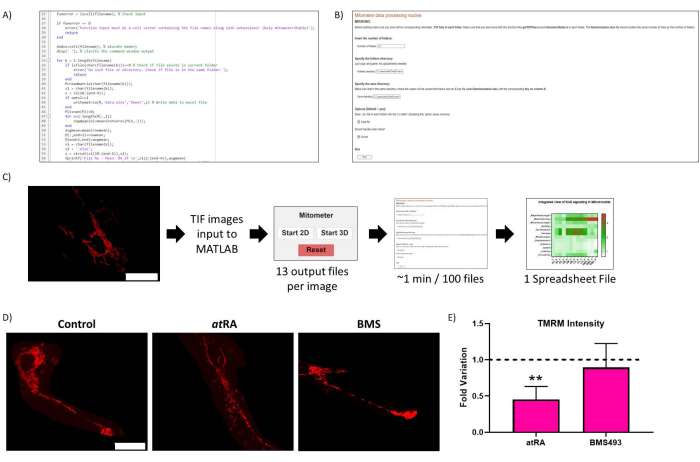
Figure 2: Routine optimization. (A) Representative code of the routine optimization. (B) Routine optimization Live-Script. (C) Routine optimization workflow. (D) Routine optimization result validation: representative image of mitochondria in untreated cells (left panel), treated with atRA (10-7 M, 72 h, middle panel), and treated with RAR antagonist BMS493 (10-7 M, 72 h, right panel), imaged after incubation with TMRM (20 nM, 45 min incubation). Scale bar = 30 µm. (E) TMRM Intensity in cell body mitochondria. Significant decrease with all-trans retinoic acid treatment (atRA, 10-7 M, 72 h), compared with control (p=0,0062), not observed when treated with RAR antagonist (BMS493, 10-7 M, 72 hours). Five cells were quantified from each of three repetitions per condition. Please click here to view a larger version of this figure.
- Image processing
- Open files in ImageJ 2.1.0 and separate position stacks by visual field: Open ImageJ and click Menu Bar | Image | Duplicate | Input slices/frame number | OK.
NOTE: To decrease repetitive inputs to the software, an ImageJ Macro was developed to facilitate duplication and saving of visual field images.
- Open files in ImageJ 2.1.0 and separate position stacks by visual field: Open ImageJ and click Menu Bar | Image | Duplicate | Input slices/frame number | OK.
- Macro protocol
- Draw a region of interest (ROI) around the cell with the free selection tool.
- Run the macro by opening ImageJ and navigating to Menu Bar | Plugins | Macros | Run Clear the background and save images macro.
NOTE: Images can be randomized in this process, storing the solution key according to optimization requirements. - Determine pixel size and voxel-depth: Open ImageJ | the Image and navigate to Menu Bar | Image | Properties.
- Alternative direct protocol
NOTE: This alternative does not use Macro to process images- Find the pixel size and voxel depth: Open ImageJ | Open Image and navigate to Menu Bar | Image | Properties.
- Draw an ROI around the cell with the free selection tool, ensuring it encompasses the whole cell throughout the 15 frames.
- Navigate to Menu Bar | Edit | Clear Outside.
- Navigate to Menu Bar | File | Save-as | Select Tiff.
- Select Saving Folder and click on Save.
NOTE: Images can be blinded/randomized manually at this point before continuing to analysis.
- Automated image analysis
- Prepare the folders for analysis files.
- Create three main folders, titled "All tracks", "Perinuclear tracks", and "Telenuclear tracks".
NOTE: These match the main automated track options. - In each main folder, create a sub-folder for each image to process, identified numerically from 1 upwards.
- Add a copy of the two routine optimization supplementary files (mitometer2table.m and getTXTfiles.m) to each image folder.
NOTE: These files help in data analysis and final format arrangement. The number of folders must match the number of elements in the randomized spreadsheet (.xlsx). After creating all numbered subfolders with supplementary files for one dataset, they can be batch-copied and pasted to the remaining datasets.
- Create three main folders, titled "All tracks", "Perinuclear tracks", and "Telenuclear tracks".
- Prepare the folders for analysis files.
- Use the MATLAB application Mitometer to analyze images.
NOTE: This protocol was run on a mitometer, installed on MATLAB R2022a. Load images in batches of 30 for optimal time-running and output balance. Maximum MAT-file size is imposed by the native file system: by default, "save" operations can create a file < 231 bytes (~2 GB); save format MAT-Files Version 7.3 can be used instead, as it allows maximum variable sizes larger than 2 GB.- Identify/alter the default MAT-file version: On the Home tab in the Medio ambiente section, click Preferences and select MATLAB | General | MAT-Files.
- Use MATLAB tool Mitometer to analyze: Open MATLAB and navigate to Menu Bar | APPS | Open Mitometer | Select Start 3D | Input data (Pixel Size (µm): 0.1395089/Time between frames (s): 2/Number z-planes: 8/Axial distance between z-planes (µm): 0.418809 | Select images to input.
- Go to Mitometer Side Menu, select Image, click on Select Highlighted, navigate to the mitometer Menu Bar | Select Tracks | Track views | select All-Tracks, Telenuclear or Perinuclear | mitometer Menu Bar | Select Analysis | Choose an element (i.e., Length) | select Save to ".txt".
NOTE: More than one parameter can be downloaded at once, if selected concomitantly. - Extract result files to the created folders (2.2.1).
- Routine optimization and data analysis
- Prepare/adapt the "Randomization.xlsx" file containing the key for image encoding.
- Insert a list of consecutive integers, from 1 upwards, in column A.
NOTE: It is advisable to keep a duplicated folder with originally named images. - Place the analytic variables in column B, composed of alphanumeric characters.
NOTE: The number of lines in the document must be consistent with the number of folders present in the main dataset. Copy and paste this "Randomization.xlsx" file into the other two main datasets.
- Insert a list of consecutive integers, from 1 upwards, in column A.
- Optimized data analysis
- Double-click "executable.mlx", input the number of folders, specify the folders directory (copy the directory from the top of the folder) | the save directory (copy the directory from the top of the folder) | the spreadsheet name in the output file and click Run.
- Perform statistical analysis as necessary.
NOTE: Optional creation of a .xlsx file in each folder with the .txt data and sound alert to the end of analysis may be selected. Live Script outputs a single spreadsheet file in a table format. In this output, columns represent analyzed parameters (e.g., "Major Axis Length"; "Intensity") and lines represent visual fields of analytic variables (e.g., "Control").
- Prepare/adapt the "Randomization.xlsx" file containing the key for image encoding.
To enhance and accelerate the analysis of output files in .txt format, a routine optimization was coded that reads data consistent with Mitometer .txt output files, with columns representing a frame and lines representing identified mitochondria. The routine optimization produces data in a single value per parameter by averaging the frames for each identified mitochondria and then averaging the results of all mitochondria per visual field. The developed routine reads files from folders numbered from 1 upwards. The Live Script Routine Optimization outputs a single spreadsheet file in a table format. In this output, columns represent analyzed parameters (e.g., "Major Axis Length"; "Intensity") and lines represent visual fields of analytic variables (e.g., "Control").
Previously published results describe mitochondrial membrane potential in the cell body of primary neuronal cultures to decrease and axonal mitochondria movement to increase after RARβ activation6.
Similar treatments were performed in differentiated neuroblastoma SH-SY5Y cells, treated with the retinoic acid receptor agonists and antagonists for 72 h (Figure 3). Collected data were plotted and analyzed using nondirectional, 2-tailed, 2-sample, equal variance Student's t-test. Comparisons were made, in the output spreadsheet file, between appropriate groups, with α = 0.05.
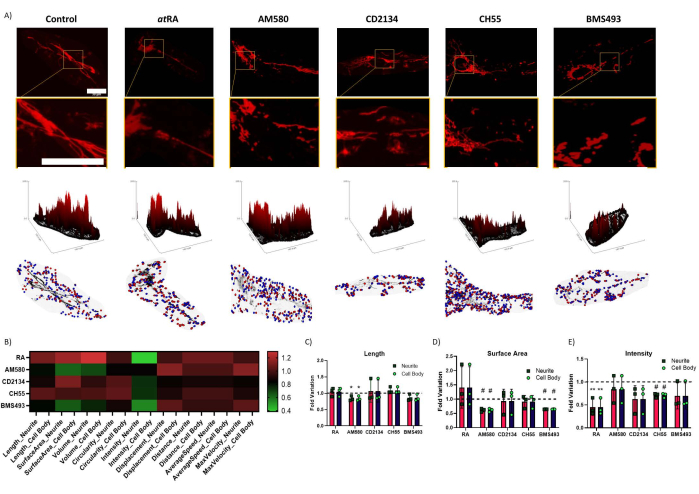
Figure 3: Regulation of mitochondrial homeostasis by isoform-specific retinoid signaling. (A) Representative image of mitochondria after treatment (top), respective surface plot (middle), and automated segmentation (bottom). Blue dots are mitochondrial initial positions; red dots are final positions. Scale bars = 30 µm. (B) Heatmap summarizing all variations found to mitochondria parameters; Significant variance was found between the cell body and neurite (two-way ANOVA, p=0.0158), with significant influence of RAR isoform-specific modulation in all mitochondria (two-way ANOVA, p=0.0082). (C) Mitochondrial Length – a significant decrease was observed in cells treated with AM580 (p=0.0179). (D) Mitochondrial surface area – significant decreases were observed in cells treated with AM580 (p=0.000406) and with BMS493 (p=3.01 × 10-8). (E) TMRM intensity, normalized for mitochondrial volume – significant decreases were found in cells treated with RA (p=0.00621) and Ch55 (p=0.000542). * p < 0,05; ** p < 0,01; # p < 0,0001. Five cells were quantified from each of three repetitions per condition. Please click here to view a larger version of this figure.
Analysis reveals that treatment of differentiated SH-SY5Y cells with RARα agonist AM580 results in a decrease in average mitochondria length and surface area; this effect was not found when treating with agonists for other isoforms other than exclusively RARα, but there was an interesting decrease of 35.42 ± 0.5% in the mitochondrial surface area following treatment with RAR pan-antagonist BMS493 (p = 3.01 x 10-8). In contrast, retinoids appear to have an opposite effect in terms of TMRM intensity, which relates to mitochondria membrane polarization11: while treatment with RARα agonist appears to not have a significant effect in TMRM intensity, treatment with RAR pan-agonist atRA results in a dramatic decrease of 54.82 ± 18.01% (p=0.00621). This decrease is also found following treatment with RARα/β agonist CH55 (28.99 ± 4.97% decrease, p=0.000542), and possibly also following treatment with RARβ specific agonist CD2314 (37.01 ± 28.96% decrease, p=0.09134). Importantly, this method differentiated between axonal mitochondria and the ones in the cell body, allowing the study of isoform-specific RAR stimulus and mitochondrial modulation.
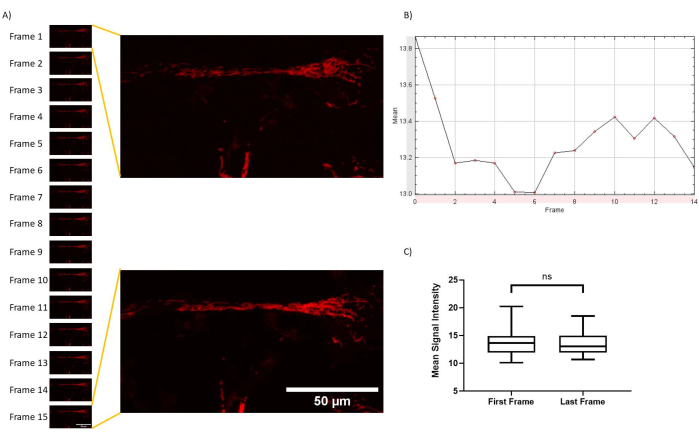
Figure 4: Minimal photobleaching throughout imaging protocol. (A) Z-stack timelapses were processed in FIJI, and z-project projections of average intensities were exported for all time points. (B) A z-axis profile of the same processing was plotted according to time. (C) Photobleaching quantification for experimental setup. No significant changes were observed (p = 0.7607; paired t-test for first and last frame mean signal intensity). Please click here to view a larger version of this figure.
