Neural tube defects (NTDs) are severe birth defects of the central nervous system caused by failures in neural tube closure (NTC) during embryonic development1. The etiology of NTDs is complex. Studies have shown that NTC involves a sequence of morphogenetic processes, including convergent extension, bending of the neural plate (e.g., apical constriction), elevating the neural fold, and finally adhesion of the neural fold. These processes are regulated by multiple molecular and genetic mechanisms2,3, and any malfunction in these processes may result in NTDs4,5,6. As mounting evidence suggests that mechanical cues also play crucial roles during NTC3,7,8,9,10,11, and relationships have been found between genes and mechanical cues12,13,14, it becomes imperative to investigate the tissue biomechanics during neurulation.
Several techniques have been developed for measuring the mechanical properties of embryonic tissues, including laser ablation (LA)15, tissue dissection and relaxation (TDR)16,17, micropipette aspiration (MA)18, Atomic Force Microscopy (AFM)-based nanoindentation19, microindenters (MI) and microplates (MP)20, micro rheology (MR) with optical/magnetic tweezers21,22,23, and droplet-based sensors24. Existing methods can measure mechanical properties at spatial resolutions ranging from subcellular to tissue scales. However, most of these methods are invasive because they require contact with the sample (e.g., MA, AFM, MI, and MP), external material injection (e.g., MR and droplet-based sensors), or tissue dissection (e.g., LA and TDR). As a result, it is challenging for existing methods to monitor the mechanical evolution of neural plate tissue in situ25. Recently, reverberant optical coherence elastography has shown promise for non-contact mechanical mapping with high spatial resolution26.
Confocal Brillouin microscopy is an emerging optical modality that enables non-contact quantification of tissue biomechanics with subcellular resolution27,28,29,30. Brillouin microscopy is based on the principle of spontaneous Brillouin light scattering, which is the interaction between the incident laser light and the acoustic wave induced by thermal fluctuations within the material. Consequently, the scattered light experiences a frequency shift, known as the Brillouin shift ωR, following the equation31:
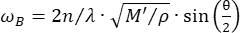 (1)
(1)
Here, 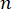 is the refractive index of the material, λ is the wavelength of the incident light, M' is the longitudinal modulus, ρ is the mass density, and θ is the angle between the incident light and the scattered light. For the same type of biological materials, the ratio of refractive index and density
is the refractive index of the material, λ is the wavelength of the incident light, M' is the longitudinal modulus, ρ is the mass density, and θ is the angle between the incident light and the scattered light. For the same type of biological materials, the ratio of refractive index and density 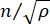 is approximately constant28,32,33,34,35,36. Thus, the Brillouin shift can be directly used to estimate relative mechanical changes in physiological processes. The feasibility of Brillouin microscopy has been validated in various biological samples29,37,38. Recently, time-lapse mechanical imaging of a live chick embryo was demonstrated by combining a Brillouin microscope with an on-stage incubation system39. This protocol provides detailed descriptions of sample preparation, experiment implementation, and data post-processing and analysis. We hope this effort will facilitate the widespread adoption of non-contact Brillouin technology for studying biomechanical regulation in embryo development and birth defects.
is approximately constant28,32,33,34,35,36. Thus, the Brillouin shift can be directly used to estimate relative mechanical changes in physiological processes. The feasibility of Brillouin microscopy has been validated in various biological samples29,37,38. Recently, time-lapse mechanical imaging of a live chick embryo was demonstrated by combining a Brillouin microscope with an on-stage incubation system39. This protocol provides detailed descriptions of sample preparation, experiment implementation, and data post-processing and analysis. We hope this effort will facilitate the widespread adoption of non-contact Brillouin technology for studying biomechanical regulation in embryo development and birth defects.
Figure 6 shows the schematic of the Brillouin microscope. The system employs a 660 nm laser as the light source. An isolator is placed right after the laser head to reject any back-reflected light, and a neutral density (ND) filter is used to adjust the laser power. A pair of lenses, L1 and L2, with focal lengths of f1 = 16 mm and f2 = 100 mm, respectively, are used to expand the laser beam. A half-wave plate (HWP) and a linear polarizer (Polarizer 1) are employed to adjust the power of the beam shining on either the sample or the calibration materials (i.e., water and methanol) after the polarized beam splitter (PBS). To focus the laser into the sample and collect backward scattered light, an objective lens (OBJ2) with a numerical aperture (NA) of 0.6 and a magnification of 40 are used. The incident light and backward scattered light are separated by the same PBS after polarization adjustment using a quarter-wave plate (QWP2). Similarly, OBJ1 and QWP1 are used to guide the laser beam to the calibration materials. The Brillouin signal is delivered by a single-mode fiber to a Brillouin spectrometer43 for analysis. Inside the spectrometer, a cylindrical lens (C1) couples the Brillouin signal into the first VIPA (virtually imaged phased array) etalon. The output of the first VIPA etalon is reshaped by another cylindrical lens (C2) and focused onto Mask 1 to reject non-scattered laser light. Then, the Brillouin signal is coupled into a second VIPA etalon by a spherical lens (SL1), reshaped by another spherical lens (SL2), and projected onto Mask 2. The resulting spectral pattern then goes through a 4-f unit for noise rejection and is projected onto an EMCCD for recording. We used a commercial microscope body (see Table of Materials). Generally, any microscope body with different brands can be used as long as it has an available port for delivering the external light beam. The on-stage incubator is a metal chamber with temperature, gas, and air flow control. The position of the sample is adjusted via the 2D motorized stage. A complementary metal-oxide semiconductor (CMOS) camera is used to acquire the bright-field image.
Figure 8 shows the time-lapse bright-field and Brillouin images of a representative chicken embryo. The embryo was extracted after 29 h of in ovo culture, and the images were taken at 29 h, 30.5 h, and 32 h with ex ovo culture, which corresponds to somite numbers of 4, 5, and 8, respectively44. In general, the embryo can be cultured in the on-stage incubator for at least 24 h. The red line in the bright-field image indicates the location for Brillouin imaging. With the acquired Brillouin images, the neural plate region was selected (highlighted by the black dotted line in Figure 8) and calculated the average Brillouin shift of this region. As shown in Figure 9, the averaged Brillouin shift of the tissue increases during neural tube closure. The longitudinal modulus is also calculated based on equation (1), where the value of = 1.3330 is estimated from literature using zebrafish embryos45, assuming the value is relatively constant for similar kinds of biological tissues28,33.

Figure 1: Schematic of the whole culture dish configuration for embryonic development. This figure shows a filter paper for attaching the embryo on the left and a 35 mm culture dish with a ring attached with a flexible film layer on the right. The cross-section schematic is also provided. Please click here to view a larger version of this figure.
Figure 2: The process of opening the eggshell above a clean 100 mm Petri dish to extract the egg into the dish. Please click here to view a larger version of this figure.

Figure 3: Filling the center well of the culture dish with thin albumen. Please click here to view a larger version of this figure.
Figure 4: Dragging away the thick albumen in the direction indicated by the red arrow, using a piece of tissue paper. Please click here to view a larger version of this figure.

Figure 5: The placement of the filter paper into the culture dish with the embryo on the upside of the filter paper. Please click here to view a larger version of this figure.
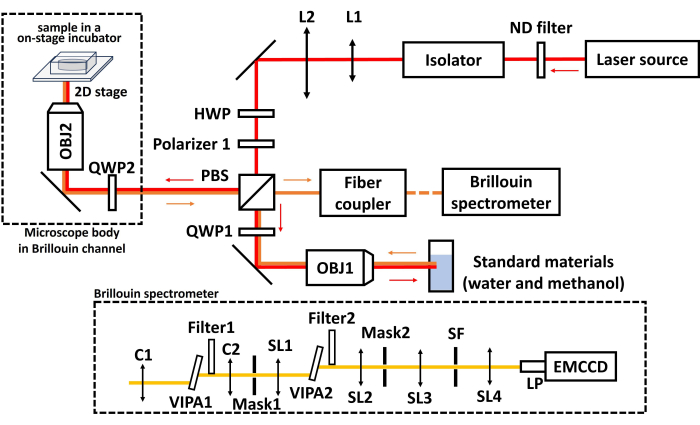
Figure 6: Schematic of the Brillouin microscope. The red arrow indicates the light path of the laser beam, and the orange arrow indicates the light path of the Brillouin signal. L1, L2, SL1-SL4: spherical lens; SF: spatial filter; C1, C2: cylindrical lens, HWP: half-wave plate; PBS: polarized beam splitter; QWP1, QWP2: quarter-wave plate; OBJ1, OBJ2: Objective lens; LP: lens pair. VIPA: virtually imaged phased array. Please click here to view a larger version of this figure.
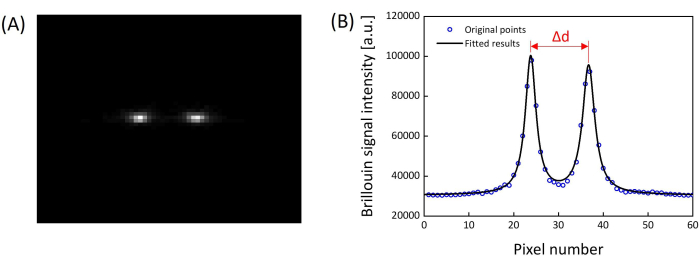
Figure 7: Representative Brillouin signal. (A) A representative Brillouin spectrum captured by the EMCCD. (B) A vertically summed Brillouin spectrum and corresponding Lorentzian fitting result. Please click here to view a larger version of this figure.
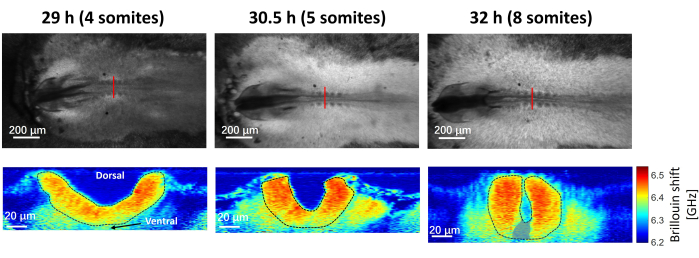
Figure 8: Bright field images and the corresponding Brillouin cross-section images of the embryo at 29 h, 30.5 h and 32 h with 4, 5 and 8 somites. Red solid lines indicate the location for Brillouin imaging, and the black dotted line indicate the neural plate region. The grey area in 32 h (8 somites) image is an artifact of curve fitting due to the weak Brillouin signal and is excluded when calculating the average shift. Please click here to view a larger version of this figure.
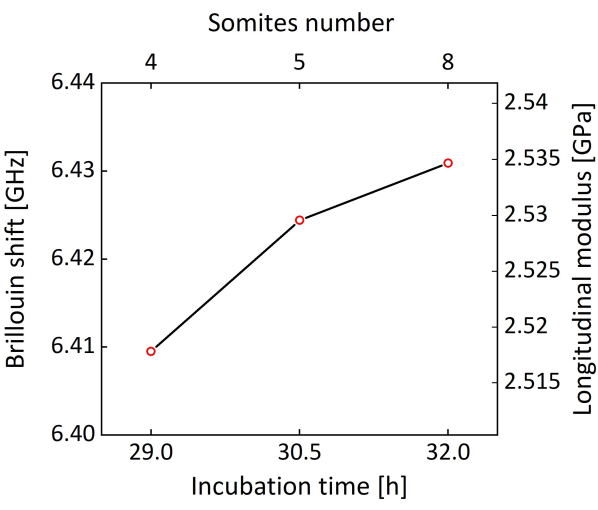
Figure 9: Brillouin shift and the estimated longitudinal modulus of the neural plate against somite number and incubation time. Please click here to view a larger version of this figure.
