In recent years, zebrafish have become highly popular model organisms for the evaluation of chemical compound effects, encompassing research areas from drug development to environmental toxicology1. As vertebrates, zebrafish share many aspects of their genetic makeup and overall physiology with humans2,3. Therefore, results obtained in this model often are directly relevant to human health. Several drug candidates currently in clinical trials have been identified in compound screens using zebrafish4.
Toxicity assessment is one major application where tests using zebrafish embryonic stages are of interest. Various Organisation for Economic Co-operation and Development (OECD) test guidelines exist for the use of zebrafish in environmental toxicity testing5,6. The small size and rapid development of zebrafish embryos make them highly suitable for screening approaches on a medium to high throughput scale1,3,4. Toxicological endpoints targeted by such screens include embryonic malformations and lethality7, endocrine disruption8, organ toxicity9, and behavioral assessments indicative of neural toxicity10,11. The behavioral assays are possible because zebrafish embryos show various types of locomotor responses to different stimuli depending on their stage of development. For example, 1 day post fertilization (dpf) embryos show spontaneous tail coiling12 and respond to a sequence of light pulses with a typical sequence of movements, the so-called photomotor response (PMR)10. After hatching, typically occurring around 48-72 hours post fertilization (hpf), the freely swimming eleutheroembryos13 gradually develop startle and escape responses to vibrational stimuli starting around 4 dpf14. These responses are characterized by a distinctive bend to the direction opposite the direction of the stimulus (the so-called C-bend or C-start), which is followed by a smaller counter bend and swimming behavior14,15,16,17. Notably, embryonic behaviors are governed by neural circuits using various neurotransmitter systems, allowing for probing chemical compound effects targeting these systems. For example, the PMR assay revealed the effects of compounds interfering with cholinergic, adrenergic, and dopaminergic signaling10, while the startle response involves cholinergic, glutamatergic, and glycinergic neurons16,18. Furthermore, compounds that damage the muscles or the neuro-muscular interface will also affect these behaviors, as will compounds toxic to the inner ear/lateral line hair cells19,20. Observing zebrafish locomotor behavior in response to a stimulus is thus a suitable means to assess not only neurotoxicity but equally ototoxicity and myotoxicity. Scoring locomotor behavior also serves as a proxy for general toxicity/lethality assessment since dead embryos do not move. Thereby, embryonic locomotion behaviors represent an integrative readout for a first-tier toxicity screening approach, which indicates lethal and neuromuscular compound effects in one setup. Given that the eleutheroembryos are already capable of metabolizing compounds, the approach may also detect the effects of metabolic transformation products7,21,22. Importantly, zebrafish embryos are not considered as protected life stage under some animal protection legislations until the stage of free feeding after 120 hpf13. Therefore, they are regarded as an alternative to animal toxicity testing.

Figure 1: Vibration startle response system setup. (A) Overview of the system. Plates with embryos exposed to the test compounds are placed on the electrodynamic transducer array ("loudspeakers"). The camera is sequentially moved by the belt-driven linear drive into the recording position above the targeted transducer. (B) Detailed view of the transducer/loudspeaker with tissue culture dish inserted on top. The plates are illuminated from below by an LED light sheet at 4000-5000 lux. An LED light next to the speaker lights up while the stimulus is given. (C) Still image of video recorded by the camera upon stimulation of the embryos. (D) Screenshot of the configuration file. (E) Screenshot of the recording software interface. Please click here to view a larger version of this figure.
Here, we describe a testing protocol for the evaluation of compound effects on the vibration startle response using an in-house build simple testing device based on vibration stimuli generated by electrodynamic transducers coupled with an automated video recording of several freely moving embryos in a tissue culture dish23. The system is modular and allows for sequential recording from several tissue culture dishes in parallel. In the setup currently used, five electrodynamic transducers provide a vibrational stimulus (500 Hz, duration 1 ms) to tissue culture dishes containing 20 embryos placed on top of them (Figure 1). The plates are illuminated from below at 4000-5000 lux with LED light sheets. An LED light next to each transducer indicates periods of stimulus application, and an oscilloscope indicates waveforms and frequency of the applied stimulus (for details, see Ref. 23). The behavior of the embryos is recorded by a high-speed camera (Table of Materials) at 1000 frames per second (fps), which is moved above the targeted speaker by a belt-driven linear drive. This recording speed is required to reliably resolve the startle response. The system provides a low-cost, individually adaptable alternative to current commercial systems. The precise workflow detailed below is currently performed in the framework of the Precision Toxicology initiative24 in order to determine suitable exposure conditions for OMICS data acquisition from zebrafish embryos treated with a selected set of toxicants.
Figure 2A shows the percentage of immotile embryos in 48 clutches of untreated wild-type embryos (AB2O2 strain). On average, 14.33% of untreated wild-type embryos do not react to the vibration stimulus. In 4 clutches, the percentage of immotile larvae reached 50%, but 75% of the clutches had a percentage of immotile larvae below 20%.
Figure 2B,C show an example of a typical calculation of a benchmark concentration/dose (BMC/BMD29,30) for compound effects on motility with the vibration startle assay workflow, as currently performed within the PrecisionTox consortium24. BMC10, BMC25, and BMC50 values correspond to the concentrations at which 10%, 25%, and 50% of embryos show immotility levels higher than the background, respectively. Only embryos that are completely immotile are included in this calculation, not those that still show partial responses, such as only a C-bend without subsequent escape swimming or only tail movements (Figure 2B). The embryos were exposed to 8 concentrations of the sodium channel inhibitor tricaine methanesulfonate, which is frequently used for fish anesthesia31. The data indicate a background level of around 25% immotility in response to the vibration stimulus. Starting at 1% tricaine, motility is reduced and then ceases above 2.5%. The KNIME workflow calculates the BMC50 as 164.9 µM, which corresponds to 1.07% tricaine and an immotility level of 75% (Figure 2C). The small 95% confidence intervals (indicated by the grey shades in the curve) indicate robust reproducibility of the motility values in this assay.
Figure 2D shows an example of a suboptimal assay run, the data of which should not be used for BMC calculations. Five E3 treated control groups with different embryos derived from the same clutch are shown (AB2O2 [wild-type strain] replicate 1-5). Only the first group shows a near normal response, showing around 25% immotility that is consistent with literature values32 and those obtained in the assay described here, as shown in Figure 2A, while all other groups show reduced and/or incomplete behavioral responses (e.g., showing only a C-bend not followed by swimming activity, or motility without a clear C-bend at the beginning). Such a response may occur when embryos do not develop properly and are in an immature state due to developmental delay, which impacts the robustness of the startle response14,33.
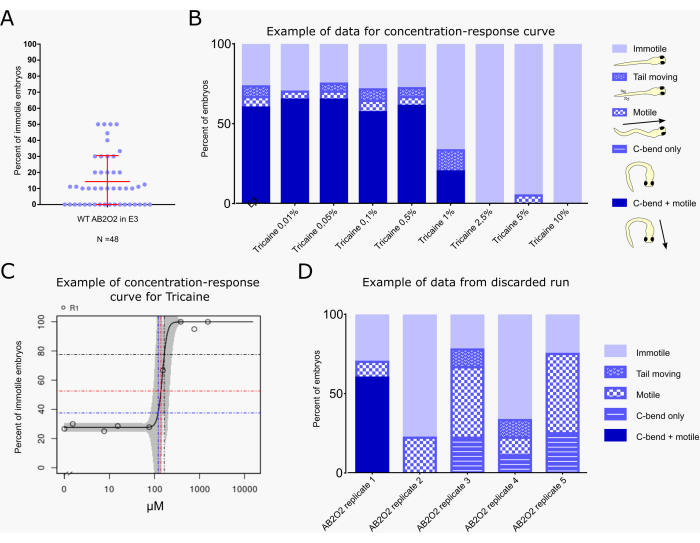
Figure 2: Example of a typical result, including benchmark dose calculation. (A) Percentage of non-responsive embryos after the sound pulse for wild-type untreated larvae for 48 clutches (n = 10 per clutch). The mean (14.33%) and standard deviation (±16.19%) are indicated in red. (B) Evaluation of startle response behavior of embryos (n=20 per condition) treated with the indicated concentration of tricaine in E3 medium or with E3 alone as a control. Behavior is classified according to the color scheme and cartoons indicated to the right of the graph, with each embryo assigned to only one of the following classes: "immotile": embryo does not show any movement; "tail moving": embryo shows tail movement, but neither C-bend nor swimming behavior; "motile": embryo shows swimming movement, but no C-bend in response to the vibrational stimulus; "C-bend only": embryo shows C-bend, but not escape swimming; "C-bend + motile": embryo shows typical C-bend behavior followed by escape swimming (the typical full startle response). The different behaviors are shown as a percentage of the total number of embryos for each treatment. (C) BMC calculation graph generated by the KNIME workflow, indicating the percentage of "immotile" embryos for each treatment concentration. Blue, red, and black lines indicate the BMC10, BMC25, and BMC50 values, i.e., the concentrations at which 10%, 25%, and 50% of embryos show immotility levels higher than the background, respectively. (D) Example of a discarded assay run. Five E3-treated control runs with different AB2O2 wild-type embryos derived from the same clutch are shown (replicate 1-5). Only replicate 1 shows a nearly normal response, while embryos of the remaining runs do not show the typical C-bend + escape swimming response. Please click here to view a larger version of this figure.
Table 1: Statistical parameters to estimate the goodness of fit and thresholds to accept determined BMC values. Please click here to download this Table.
Table 2: Properties of a selection of vibrational startle response assay systems. Please click here to download this Table.
Supplementary File 1: Excel template for configuration file. Please click here to download this File.
Supplementary File 2: KNIME input template with an example data set. Please click here to download this File.
Supplementary File 3: KNIME output file example. Please click here to download this File.
