Clinical scores assessment
As shown in Figure 1, the mice in the control group did not exhibit any clinical symptoms. The mice in the EAE group, which were immunized with MOG35-55, showed tail paralysis approximately 12 days after immunization. By day 16, the symptoms reached complete hind limb paralysis (defined as peak stage, Peak). After that, the symptoms gradually were remitted. Mice's clinical symptoms were aggravated at around day 25 and reached complete hind limb paralysis at approximately day 30 (defined as a relapsing stage, RR). Notably, the symptoms lasted longer than the previous time, which developed a classical relapsing-remitting feature. The principal manifestation of relapsing-remitting EAE mice experience different stages of outbreak, remission, and relapse during the progression of the disease, which have a certain periodicity and fluctuation.
Histological analysis
The control group mice displayed intact brain tissue structure with evenly distributed and clearly defined blood vessels, without any inflammatory cell infiltration. (Figure 2A). At the peak stage of the EAE group mice, a significant infiltration of inflammatory cells was observed in the brain (Figure 2B), forming cuff-like changes around blood vessels (indicated by red arrow). Similarly, in EAE group mice at the RR stage, there was evident infiltration of inflammatory cells (Figure 2C), with a predominant aggregation around blood vessels (indicated by red arrow).
LFB staining analysis
In the control group, the myelin sheath of the spinal white matter appeared blue, indicating that its structure was intact and tightly packed (Figure 3A). At the peak stage of the EAE group, the spinal white matter exhibited a light blue or white color; the structure appeared to be loose and discontinuous, with a significant presence of vacuolar degenerations indicated by a red arrow (Figure 3B). In the RR stage of the EAE model group, extensive loss of white matter myelin in the spinal cord was observed. The structure appeared to be loose, and a significant number of vacuolar degenerations could be observed (indicated by red arrows; Figure 3C).
Flow cytometry assay
The results depicted in Figure 4A,B demonstrated a significant increase in the proportion of CD4+CD69+ Teff cells in EAE mice at the peak stage when compared to the control group (P < 0.0001). Additionally, the proportion of these cells also increased during the RR stage, although the difference was not statistically significant (P > 0.05). Figure 4C,D revealed a notable rise in the proportion of CD4+CD44+CD62L– Tem cells in EAE mice at both the peak and RR stages, as compared to the control group (P < 0.0001).
Cytokine measured by cytometric bead array
The results depicted in Figure 5 demonstrate that, when compared to the control group, the levels of IL-4 (P < 0.0001), IL-6 (P < 0.001), IFN-γ (P < 0.0001), and TNF-α (P < 0.0001) were significantly elevated in the EAE group at the peak stage. While the expression of IL-17A also increased, the difference was not statistically significant (P > 0.05). Moreover, the level of IL-2 exhibited no change during the peak stage of EAE mice (P > 0.05); however, it significantly increased during the RR stage (P < 0.01). Furthermore, both TNF-α (P < 0.001) and IL-17A (P < 0.01) expressions in the RR stage were significantly higher than those observed in the control group. Conversely, the expression of IL-10 in the EAE group was significantly lower than that in the control group, both at the peak and RR stages (P < 0.0001).
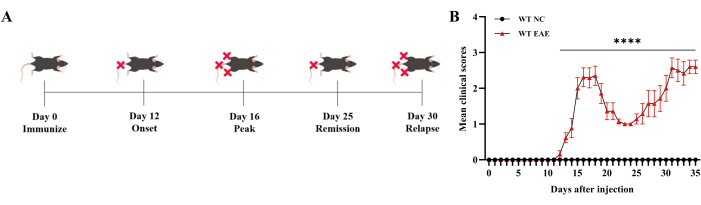
Figure 1: Clinical score assessment. The clinical score was assessed daily and shows that mice developed a classical relapsing-remitting EAE immunized by MOG35-55. (A) The diagram of symptom progression of the EAE mice. (B) The assessment of EAE mice through mean clinical scores. Data are expressed as mean ± SEM, n = 6. The data were analyzed using one-way ANOVA with post-hoc comparisons to assess statistically significant differences. * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001. Please click here to view a larger version of this figure.
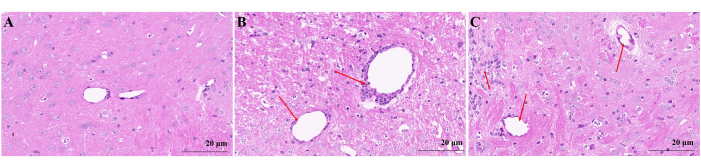
Figure 2: Microscopic map of brain H&E staining. (A) Mice brain in the control group. (B) EAE mice brain in peak stage. Inflammatory cells were infiltrated in the brain at the peak stage in EAE group mice and distributed around blood vessels to form cuff-like change (red arrow). (C) EAE mice brain in RR stage. Inflammatory cells in EAE group mice of RR stage aggregated around blood vessels (red arrow). Please click here to view a larger version of this figure.
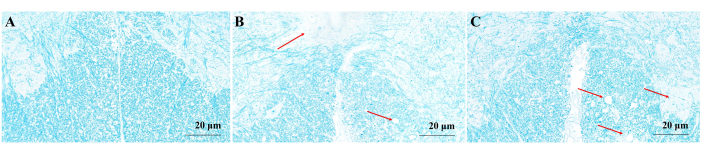
Figure 3: Microscopic map of myelin sheath LFB staining. (A) Mice myelin sheath in the control group. (B) EAE mice myelin sheath in peak stage. The spinal white matter was light blue or white, the structure was loose and discontinuous, and a large number of vacuolar degenerations appeared (red arrow). (C) EAE mice myelin sheath in RR stage. The white matter myelin of the spinal cord was widely lost, the structure was loose, and a large number of vacuolar degenerations appeared (red arrow). Please click here to view a larger version of this figure.
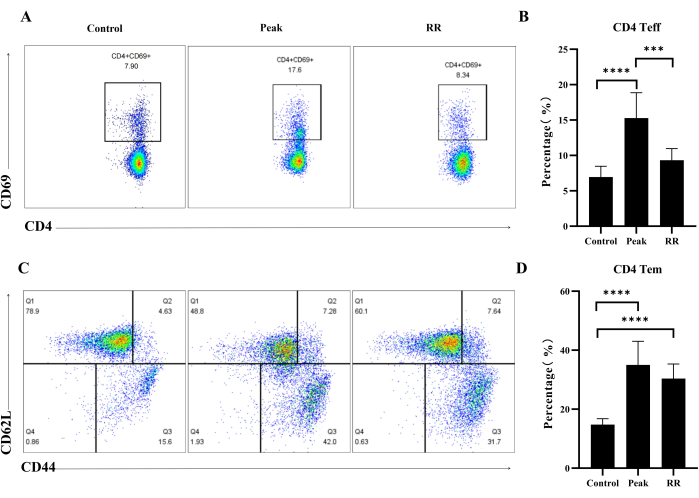
Figure 4: Immunization by MOG35-55 induced the up-regulation of CD4 Teff and Tem cells proportion in both peak and RR stage in EAE mice. (A) Illustrative flow plot displaying the immunostaining of CD4+CD69+ cells. (B) Percentage of CD4+CD69+ Teff cells. (C) Illustrative flow plot displaying the immunostaining of CD44+CD62L+ cells (gate on CD4 T cells). (D) Percentage of CD44+CD62L+ Tm cells. Data are expressed as mean ± SEM, n = 5. The data were analyzed using one-way ANOVA with post-hoc comparisons to assess statistically significant differences. * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001. Please click here to view a larger version of this figure.
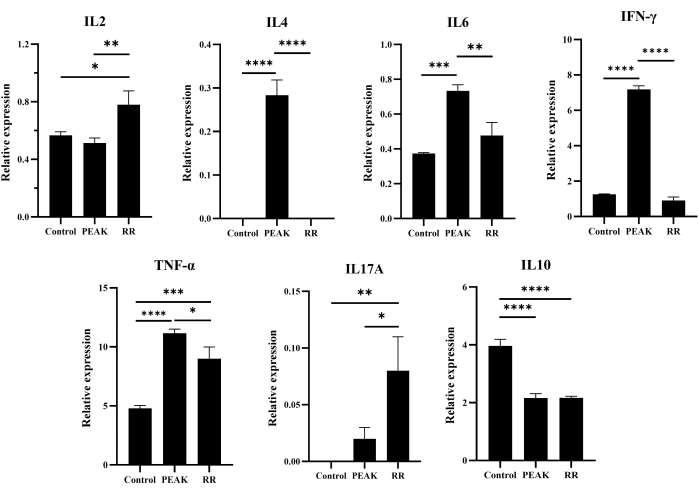
Figure 5: Level of cytokines in mice brain analyzed by cytometric bead array. Data are expressed as mean ± SEM, n = 3. The data were analyzed using one-way ANOVA with post-hoc comparisons to assess statistically significant differences. * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001. Please click here to view a larger version of this figure.
| Tube label | Concentration(pg/mL) | Cytokine Standard dilution |
| 1 | 0 (negative control) | no standard dilution(Assay Diluent only) |
| 2 | 20 | 1:256 |
| 3 | 40 | 1:128 |
| 4 | 80 | 1:64 |
| 5 | 156 | 1:32 |
| 6 | 312.6 | 1:16 |
| 7 | 625 | 1:8 |
| 8 | 1250 | 1:4 |
| 9 | 2500 | 1:2 |
| 10 | 5000 | Top standard |
Table 1: List of cytokine standard dilutions to the control tubes.
