First, we evaluated the cells grown in a 24-well plate for 7 days using brightfield imaging. We identified small clusters of cells assembling into organoids during the week, as seen in Figure 4. A controlled scan method can follow the mobility of the cells and the organoids between different days. In general, we looked at the evolution of the morphology of the cells during the whole week. SW1222-derived organoids should have a round morphology and a light appearance. A darker appearance indicates an undesired higher cell density, as seen in some of the colonies cultured in Peptide P, day 7 (Figure 4).
Figure 5 shows the results of the cell viability (Figure 5A) and proliferation assays (Figure 5B), which point towards the amount of dead and live cells and the metabolic activity, respectively. Here, we used a calcein and ethidium homodimer-based viability assay and noticed that these cells showed a small population of dead cells by day 7 when cultured in Matrigel. This phenomenon was also present in all peptide-based hydrogels but not in the 2D culture. Proliferation results showed a total increase in metabolic activity between day 1 and day 7. Moreover, we found slight decreases in activity between day 4 and day 7 for several conditions. This observation correlates to the increase in the dead cell population seen in the viability assay (Figure 5A).
For a preliminary evaluation of the interaction between the cells and the biofunctionalization of the peptide hydrogels, we performed an adhesion assay based on mechanical stress to induce cell detachment. As can be observed in Figure 6, the use of different concentrations of the biofunctional peptide P2 significantly affected the number of cells retained before and after the detachment process. Here, we used a peptide that contains only the biofunctional motif of peptide P2 as a positive control. We also used the non-biofunctional peptide P as a negative control. As can be observed in the graph, cells seeded in peptide P had a relatively low initial adhesion as well as a low retention value. Moreover, the change in the ratio of the biofunctional peptide P2 was correlated to the amount of cells retained after the stress. Hence, using this assay, we can conclude that the P2 peptide affected cell adhesion that may or may not be driven by the biofunctional motif but was correlated with the amount of biofunctional peptide present in the mixture.
Then, we investigated the effect of the matrices in the fabricated organoids. We evaluated the circularity of the colony and the relative size of the lumen with respect to the size of the colony by performing shape analysis of cytoskeleton-stained samples cultured for 4 days. The data points had a particular distribution along the x- and y-axes, and each condition had a centroid with a particular value for circularity and relative lumen area. Figure 7 shows that the cells cultured in 2D had a very high variation in colony circularity. In contrast, the cells cultured in Matrigel had a more extensive distribution for the relative size of the lumen.
Interestingly, the centroid values for all peptide-containing hydrogels had a very similar position. However, the distance between data points showed different distribution profiles. For instance, the P3 peptide had a smaller distribution among the three peptide hydrogels, while the data points from the non-biofunctional peptide P appeared to have a larger distribution. These data suggest that both hydrogels with P1 and P3 have the potential to be optimized for organoid culture.
Finally, we evaluated the expression of specific markers using immunocytochemistry on a sample stained for the apical markers ZO-1 and ezrin. These markers allowed for the evaluation of the polarization of the cells within each organoid. We also evaluated pan cadherin markers to visualize cell-cell junctions and a cytoskeleton-specific marker (phalloidin) to visualize the lumen. Figure 8A shows the organoids found in non-biofunctional peptide P, biofunctionalized peptide P1, and Matrigel on day 4 and day 7. ZO-1 and ezrin markers should be localized within the apical and basolateral membranes of the organoid; proper cell polarization will allow us to observe the expression of both molecules by day 4 near the lumen area, otherwise invisible. Indeed, as seen in Figure 8, ZO-1 and ezrin markers were located in the apical membranes of the organoids cultured in Matrigel. Interestingly, ezrin markers were barely visible in the non-biofunctional peptide P. From this, we infer that the inefficient polarization of the cells resulted in a low expression of these molecules in peptide P.
Moreover, the dispersion of the nuclei tended to be irregular in the peptide hydrogels, whereas the organoids embedded in Matrigel formed a single, asymmetrical layer of nuclei. This description matches the ideal morphology for a colorectal organoid; however, although the orientation of the nuclei was not completely asymmetrical in the peptide hydrogels, denser colonies with multilayers of nuclei were found mainly in the non-biofunctional peptide P. In contrast, cell-cell junctions must be localized perpendicular to the apical and basolateral membranes (Figure 8B). This marker allowed us to observe the edges of all cells located within the colony. When grown in Matrigel, cells had an almost perfectly radial distribution of pan-cadherin (cell-cell junctions) by day 7. In contrast, non-polarized colonies, such as the ones in peptide P, presented a disordered arrangement of cells, and expression of cell-cell junctions near the basal and apical membrane.
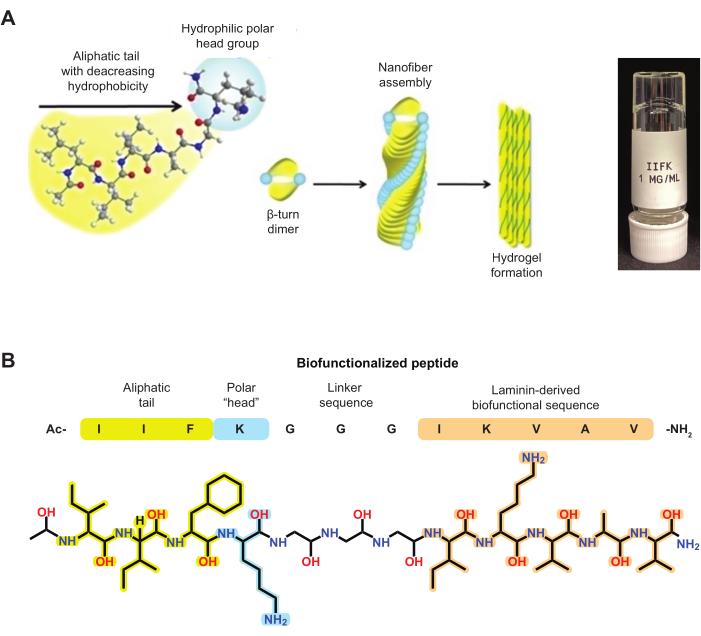
Figure 1: Ultrashort self-assembling peptides forming nanofibers. (A) Depiction of the self-assembling process for aliphatic ultrashort peptides. (B) Chemical structure of ultrashort SAP IIFK biofunctionalized with the IKVAV ligand. This figure was modified from Loo et al.4. Please click here to view a larger version of this figure.

Figure 2: Structure and properties of peptides. (A) The chemical structures of the parent peptide IIFK and the biofunctional peptide P2 are illustrated. (B) The inverted vial test results reveal critical gelation concentration and gelation time differences. P2 forms a translucent gel with a higher CGC than the parent peptide IIFK. Notably, the IIFK:P2 (1:1) mixture exhibits an even higher CGC and longer gelation time than either peptide alone. (C) Scanning electron micrographs provide visual insights into the structural characteristics of IIFK, P2, and the IIFK:P2 (1:1) mixture. (D) The atomic force microscopy height topography images depict the gelated peptides at a concentration of 8 mM, offering a detailed view of their surface features. (E) Circular dichroism spectroscopy reveals the conformational changes of the IIFK:P2 mixture at various concentrations, shedding light on their secondary structure alterations. (F) Rheological analyses measure the stiffness of the gelated peptides at different concentrations, providing valuable insights into their mechanical properties. This figure is reproduced from Perez-Pedroza et al.25. Scale bars = 1 µm (C), 200 nm (D). Abbreviation: CGC = critical gelation concentration. Please click here to view a larger version of this figure.
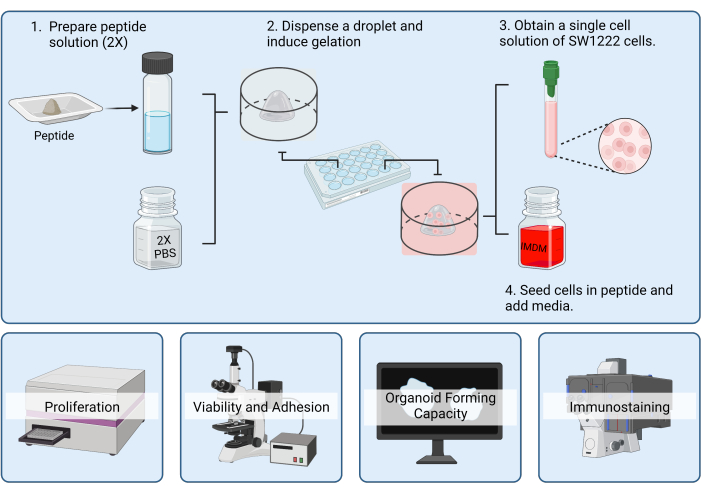
Figure 3: Diagram depicting peptide preparation and cell seeding for organoid fabrication and characterization by viability, adhesion, organoid forming capacity, and immunostaining in peptide hydrogels. Please click here to view a larger version of this figure.
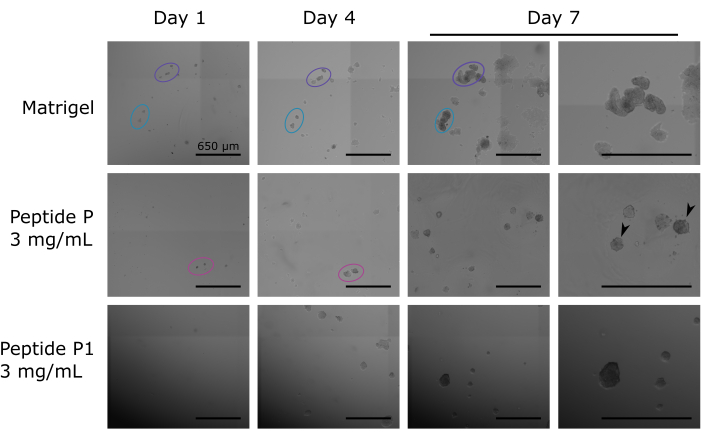
Figure 4: Brightfield microscopy images captured from SW1222 cells cultivated within peptide hydrogels labeled as P and P1 compared with Matrigel control. Interestingly, organoids cultured in the peptide hydrogels manifest a predominantly spherical morphology by the seventh day. In contrast, those situated within the Matrigel seem to be exhibiting a lowered cell density, which can be told by their bright body, compared to the darkened body of the organoids culture in SAP (black arrowheads). Nevertheless, they exhibit an apparent clustering between adjacent colonies. Furthermore, it becomes evident that the migratory capacity of cells within the Matrigel environment is comparatively diminished. Scale bars = 650 µm. Please click here to view a larger version of this figure.
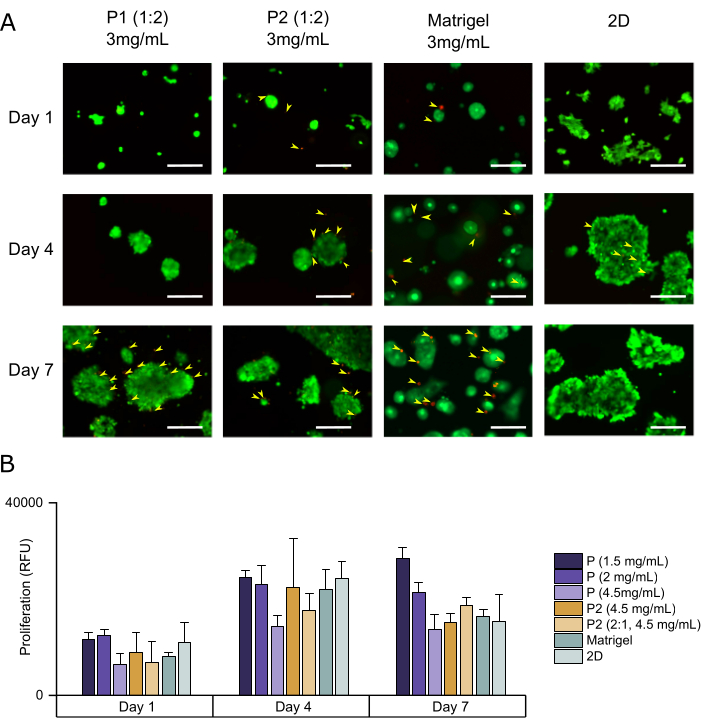
Figure 5: Viability and proliferation assays of SW1222 cells in biofunctionalized SAP at various concentrations. (A) Viability assessment of cells cultured in biofunctional peptide P1 and P2, compared to Matrigel and 2D. (B) Proliferation assessment of cell culture in non-biofunctional peptide P1 and biofunctional peptide P2, compared to Matrigel and 2D controls. Here, we see a behavior similar to that of the cells growing in Matrigel for the live/dead assay. A slight increase in dead cells (yellow arrowheads) is found in several conditions, mainly Matrigel. For proliferation, the trends vary depending on the type and concentration of the peptide, but generally, there is a positive trend. Scale bars = 250 µm. This figure was adapted from Perez-Pedroza et al.25. Please click here to view a larger version of this figure.
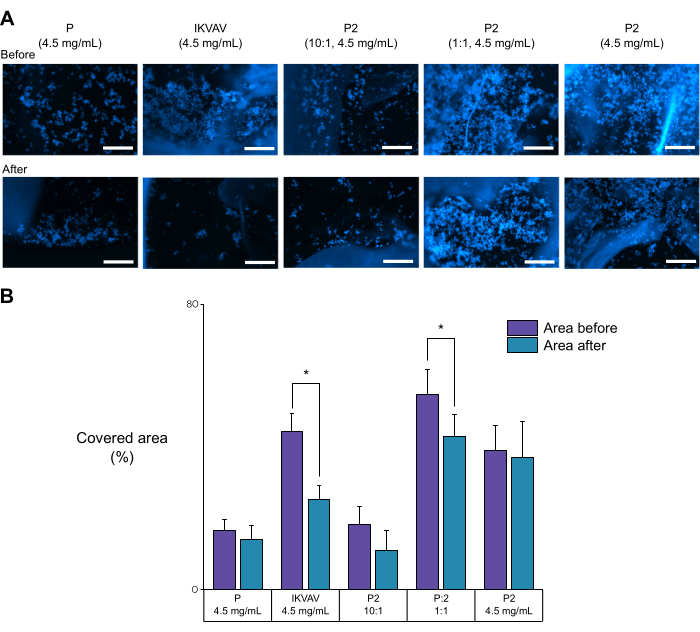
Figure 6: Effects of the peptide on the number of cells detached. (A) Fluorescence images of cells stained with DAPI before and after mechanical stress. Here, we use peptide P, modified peptide P2, and the IKVAV motif. (B) Change in cell adhesion was determined by measuring the covered area in ImageJ. The comparative analysis revealed that the biofunctionalized peptide P2 had a heightened retention capacity compared to the non-biofunctional peptide P. Scale bars = 100 µm. This figure was adapted from Perez-Pedroza, et al.25. Please click here to view a larger version of this figure.
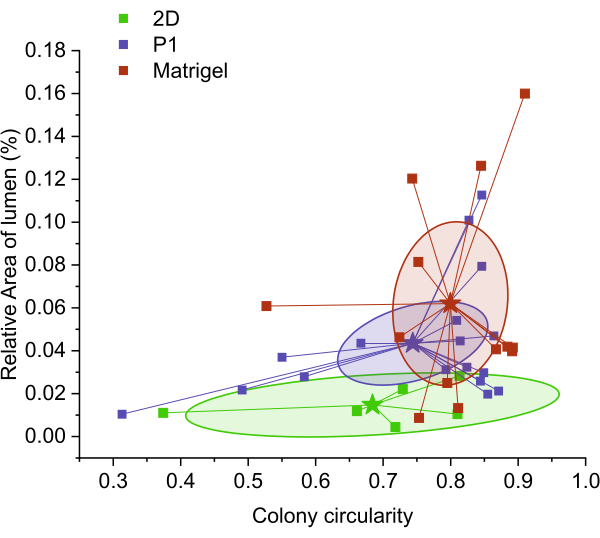
Figure 7: Evaluation of organoid forming capacity by centroid calculation of relative area of the lumen versus circularity. The centroid and dispersion of the data varied among treatments. As expected, the worst-performing condition was that of the 2D measurements, which expanded in an extensive range of circularity values and had the lowest relative area of the lumen. Matrigel had not only the highest circularity but also the biggest lumen. The peptide conditions varied slightly in both values, but the P3 condition held the most negligible variation of the data. Please click here to view a larger version of this figure.
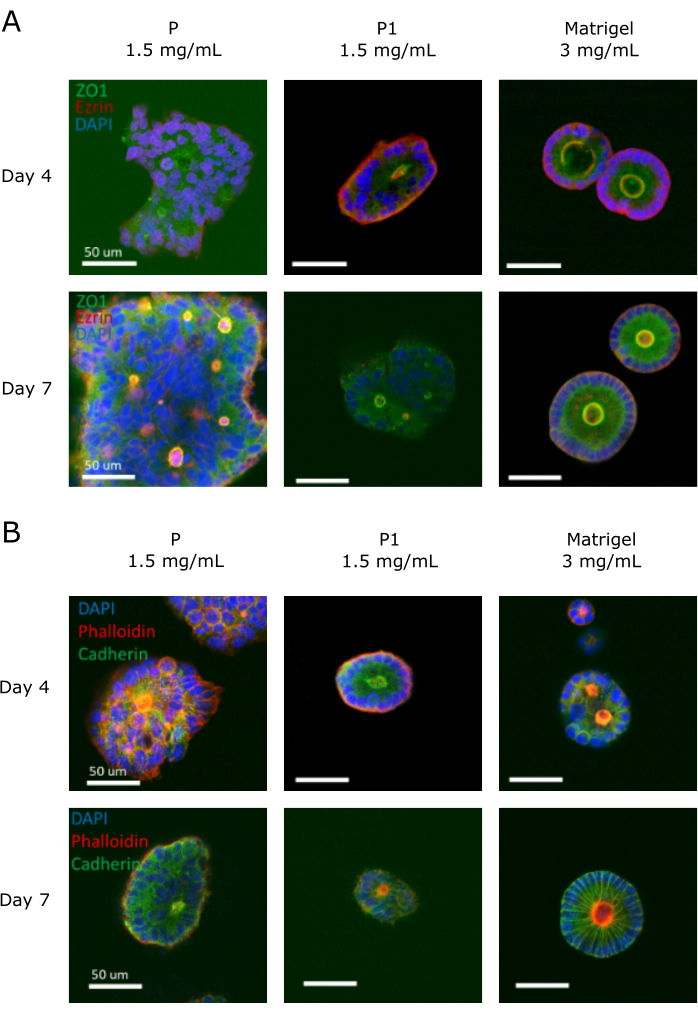
Figure 8: Confocal microscopy images of organoids at day 4 and day 7 for various markers. (A) ZO-1 (green), ezrin (red), and DAPI (blue) markers are used to find the apical distribution of the cells within the organoids. As seen in the Matrigel control, these molecules are typically localized in the apical and basolateral membranes. Non-biofunctional peptide P shows expression of these molecules only by day 7. (B) Cell-cell junction markers stained with pan-cadherin (green) antibodies should be perpendicular to the basolateral and apical membranes of the organoids. Cell nuclei (blue) and cytoskeleton (red) are used as counterstaining. The non-biofunctional peptide P induced cell-cell junction expression in the basolateral membrane, contrary to the other two treatments (P3 and Matrigel). Scale bars = 100 µm. Please click here to view a larger version of this figure.
