Using this culture protocol, primary epicardial cells can be isolated with high purity for downstream applications. The cultured cells are able to undergo EMT, migrate and differentiate just as epicardial cells do in vivo.
To determine the purity of our primary epicardial cell culture, we analyzed the epicardial explants generated from Sema3dGFPCre/+;R26Tom/+ embryos. Sema3d is expressed in epicardial cells during early cardiac development, thus epicardial cells derived from these embryos will be RFP labelled. We isolated ventricles from Sema3dGFPCre/+;R26Tom/+ embryos for epicardial explants. After 48-72 hr, we were able to see a monolayer of epicardial cells. Superimposing the RFP immunofluorescence and brightfield images showed that a majority of the cells that migrated from the ventricles are of epicardial origin (Figure 1A–C). We further validated our results by isolating RNA from primary epicardial cells and performing qPCR which showed a strong expression of epicardial specific genes (Tbx18 and Wt1) as opposed to a cardiomyocyte marker gene (Nkx2.5) (Figure 1C).
Next we examined the ability of the cells to undergo EMT by analyzing the cell polarity and cell-cell contact of epicardial cells. EMT is a biological process that allows an epithelial cell to lose its cell polarity and cell-cell contact in order to transform into a migratory mesenchymal cell. The first step of EMT is the detachment of cell-cell contacts. We immunostained epicardial cells with ZO-1 (also known as Tjp1, Tight junction protein 1) which showed localization of ZO-1 to the plasma membrane indicating that cells have yet to undergo EMT (Figure 2A). Next we performed immunostaining for α-Tubulin (Figure 2B) so as to observe the organization of the microtubules in epicardial explants, which showed a polarized alignment that facilitates directional migration of epicardial cells. In addition, to determine the differentiation potential of the epicardial cells, we cultured them for 6 days in a differentiation media containing recombinant TGF-β1. Immunostaining for SMA demonstrated successful differentiation of epicardial cells into smooth muscle cells (Figure 2C).
Lastly, to analyze epicardium-derived cell migration and epicardial EMT, we also performed a collagen gel invasion assay. Epicardium-derived cells were visualized by phalloidin immunostaining. Successful migration of epicardium-derived cells can be seen all around the explant (Figure 3A). To determine the depth of cell penetration into the collagen matrix, 3D-reconstruction was generated from confocal images. Penetration of epicardium-derived cells can be seen in z-stacks (Figure 3B).
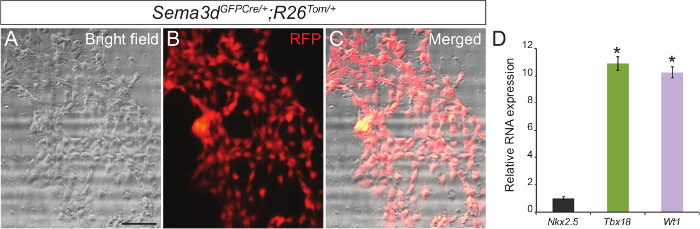
Figure 1: Primary Culture of Epicardial Cells from Embryonic Mouse Hearts. Representative brightfield (A) and RFP immunofluorescence (B) images of primary epicardial cells generated from E12.5 Sema3dGFPCre/+;R26Tom/+ hearts. Merged brightfield and immunofluorescence image (C). Relative mRNA expression levels of epicardial markers (Tbx18 and Wt1) and cardiomyocyte marker (Nkx2.5) in primary epicardial cells (D). The results were normalized to Gapdh expression, and the relative expression levels are given as a fold difference compared to Nkx2.5 expression (n = 3). The data are presented as the mean ± SD. Scale bar = 200 µm. *, p <0.05. Please click here to view a larger version of this figure.
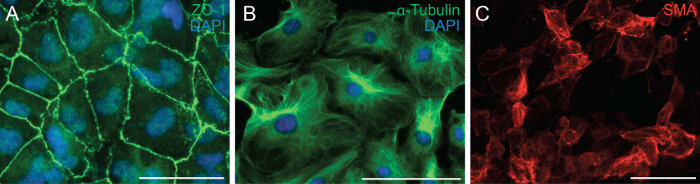
Figure 2: Primary Epicardial Cells Retain their Ability to Undergo EMT and Differentiation. Immunostaining on epicardial explants for ZO-1 (A, green) or α-Tubulin (B, green). DAPI was used to visualize nuclei (blue). Epicardial cells were differentiated into smooth muscle cells for 6 days and stained with α-smooth muscle actin (SMA) (C, red). Scale bar = 100 µm. Please click here to view a larger version of this figure.
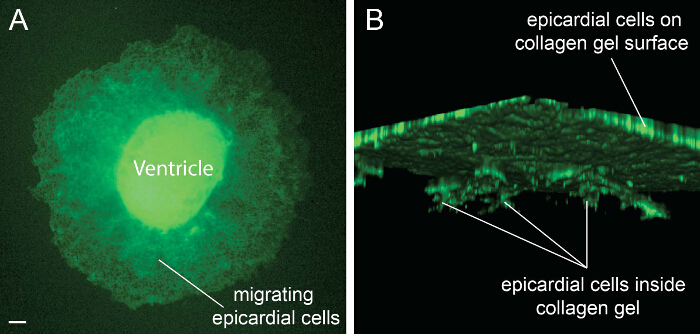
Figure 3: Epicardial Cell Migration on the Collagen Gel Surface. Ventricles from E12.5 embryos were cultured on a three-dimensional collagen gel matrix and stained with Alexa Fluor 488-phalloidin to visualize epicardium-derived cell migration and penetration (A). Confocal images were used to construct a three-dimensional image to determine cell penetration along the z-axes (B). Scale bar = 50 µm. Please click here to view a larger version of this figure.
